 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
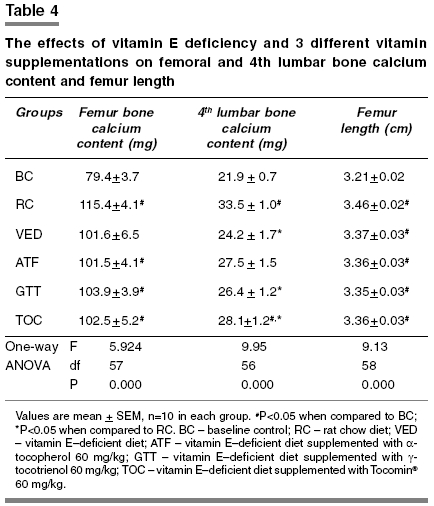
Indian Journal of Pharmacology, Vol. 37, No. 5, September-October, 2005, pp. 309-314 Research Paper Gammα-tocotrienol is required for normal vitamin D metabolism in female rats Norazlina M, Ng FW, Ima-Nirwana S Dept of Pharmacology, Faculty of Medicine, Universiti Kebangsaan Malaysia, Kuala Lumpur Date of Submission: 24-Feb-2005 Code Number: ph05080 Abstract Objective: To study the effects of vitamin E deficiency and supplementation of vitamin D and bone metabolism in female Sprague-Dawley rats.Materials and Methods: Rats weighing between 200 and 250 g were divided into six groups, that is, rats fed on (a) normal rat chow diet (RC), (b) vitamin E-deficient diet (VED) (c) VED diet supplemented with 60 mg/kg α-tocopherol acetate (ATF), (d) VED diet supplemented with 60 mg/kg γ-tocotrienol (GTT), (e) VED diet supplemented with 60 mg/kg Tocomin ® (TOC) and (f) baseline control group which was killed without treatment (BC). Treatment was given for 2 months. Serum 1,25-dihydroxyvitamin D3, serum total calcium, urine calcium, left femur, and fourth lumbar vertebra calcium content and left femur length were measured. Results: In the VED and ATF groups, activation of vitamin D to 1,25-dihydroxyvitamin D3 was inhibited and calcium reabsorption in the kidneys were increased. Both the effects seen in ATF and GTT groups were observed in the TOC group. The GTT group was protected from the effects of the vitamin E-deficient diet. Calcium content of the fourth lumbar vertebra was also decreased by vitamin E- deficiency, which was not reversed by vitamin E supplementation. Conclusion: γ-tocotrienol and not α-tocopherol protects vitamin D metabolism and calcium homeostasis from the effects of vitamin E-deficiency. Keywords: Calcium homeostasis, α-tocopherol, vitamin E-deficiency Introduction Vitamin D and its metabolites are among the factors that are important in regulating bone metabolism. Vitamin D3 is a prohormone, which is hydroxylated in the liver and in the kidney to the active metabolite 1,25-dihydroxyvitamin D3 and 24,25-dihydroxyvitamin D3. In a previous study, supplementation with a 1, 25-dihydroxyvitamin D3 analogue was able to increase bone mineral density in osteoporotic women.[1] The most potent vitamin D metabolite is 1,25-dihydroxyvitamin D3 (calcitriol), which stimulates calcium and phosphate transport from the intestine and calcium reabsorption from bone.[2] 1,25-dihydroxyvitamin D3 has also been reported to act directly on circulating osteoclast precursors to influence osteoclast differentiation.[3] Vitamin E, a lipid-soluble vitamin with antioxidant properties has an important role in protecting biological systems. There are two types of vitamin E; tocopherol and tocotrienol, and there are 4 isomers for each type: alpha (α), beta (β), gamma (γ) and delta (δ). Tocopherol has been shown to prevent lipid peroxidation[4] and heart disease in postmenopausal women.[5] Tocotrienol was shown to reduce carcinogenesis,[6] and to inhibit the production of HMG-CoA reductase - the enzyme responsible for cholesterol synthesis.[7] Palm oil is the major source of vitamin E, mainly the tocotrienol type. Vitamin E-rich extract from palm oil protected the bone against the toxic effects of the oxidizing agent ferric nitrilotriacetate,[8] reduced bone resorption, and increased the survival rate of thyrotoxic rats[9] and prevented the decline in bone mineral density, and bone calcium loss seen in orchidectomized rats.[10] Palm vitamin E (60 mg/kg) was comparable to α-tocopherol in maintaining bone mineral density and bone calcium content in ovariectomized rats.[11] Tocotrienols and δ-tocopherol were able to induce apoptosis in human mammary gland tumor cells.[12] However, a few studies found that tocotrienol is superior compared to tocopherol. Alphα-tocotrienol was able to protect against glutamate-induced neuron cell death compared to α-tocopherol.[13] Palm vitamin E improved bone calcium content in vitamin E- deficient rats compared to α-tocopherol.[14] Vitamin E deficiency impaired calcification[15] and retarded bone growth[16] in normal and ovariectomized rats. Its mechanism is unclear. One hypothesis is that vitamin E deficiency may influence the hormones regulating bone metabolism (e.g., vitamin D metabolism). Other studies have shown that the presence of vitamin E is crucial in vitamin D metabolism and calcium absorption from the intestines in which they found that vitamin E deficiency reduced calcium absorption from the intestines[17] and inhibited the conversion of vitamin D to its active metabolites.[18] To further examine this hypothesis, this study was carried out, in which the effects of a vitamin E-deficient diet on vitamin D metabolism and bone calcium homeostasis were determined. The effects of supplementation with three different types of vitamin E on vitamin D metabolism were also studied. Materials and Methods Animals and treatment Diets Alpha-tocopherol (60 mg/kg, body weight) was prepared by mixing 3 g of a-tocopherol in 47 g olive oil. The same procedure was applied in preparing γ-tocotrienol and Tocomin ® (60 mg/kg, b.w.). 0.1 mL/100 g rat weight of the respective solutions was given by oral gavage 6 days a week for 2 months. Bone and blood samples collection At the end of the treatment period, the rats were anesthetized and then sacrificed. Left femoral and 4th lumbar bones were dissected out and cleansed of all soft tissues. Femur length was measured and the bones were then stored at -700C for the measurement of bone calcium content. Urine collection Serum 1,25-dihydroxyvitamin D3 level Serum and urine calcium levels Bone calcium content This study was approved by the University′s Research and Animal Ethics Committees and was carried out in accordance with the guidelines stated by the committees. Analyses of data Results Serum 1,25-dihydroxyvitamin D3 level Total serum calcium level Urine calcium level Bone calcium content Femur length Discussion Our previous study showed that vitamin E deficiency caused hypocalcemia in rats as early as the first month of treatment;[19] however, calcium levels were almost back to normal in the second month. We also found that secretion of parathyroid hormone was increased in the second month and it stimulated bone resorption and corrected hypocalcemia. On the basis of our findings in the previous study, we fixed the treatment duration for the present study at 2 months because biochemical changes are likely to occur within this period. In the present study, no significant difference in serum calcium level can be seen between the treatment groups. As per the results of our previous study, hypocalcaemia, which may have occurred, was probably corrected by the second month. Studies have shown that vitamin E deficiency-induced disturbances in calcium transport from the intestine[17] were due to the inhibition of the conversion of vitamin D to its active metabolites.[18] These researchers also found that the rats showed a decreased level of 1,25-dihydroxyvitamin D3. Although the study by Sergeev et al . (1990)[18] failed to show an increase in parathyroid hormone level, our study showed the opposite.[19] However, the treatment period and the time point at which parathyroid hormone levels were measured might have been different between these two studies. Our present study measured total serum calcium, which might not accurately reflect the ionized calcium level. Vitamin E deficiency might affect levels of plasma proteins, although there is no evidence to support this; therefore in future studies, measurement of ionized calcium might be more meaningful. Measurement of phosphorus levels should also be considered because phosphorus is another major mineral which contributes to bone density and bone growth. In the present study, no significant difference in serum 1,25-dihydroxyvitamin D3 level was observed between all groups at baseline and upon completion of the study. However, upon within group comparison, the ATF and TOC groups showed a significant decrease in serum 1,25-dihydroxyvitamin D3 levels after treatment compared to baseline values. Low levels of 1,25-dihydroxyvitamin D3 in ATF and TOC groups suggests that vitamin E deficiency inhibit the activation of vitamin D, and supplementing with ATF or TOC did not overcome the inhibition. The observation that no significant change in 1,25-dihydroxyvitamin D3 levels before and after treatment for the VED group suggests that after 2 months, some compensatory mechanisms enabled the metabolism of vitamin D to return to normal. The inability of the ATF or TOC groups to normalize the 1, 25-dihydroxyvitamin D3 levels after 2 months suggest inhibition of the compensatory mechanism that counteract the effects of vitamin E deficiency. The compensatory mechanism, which is responsible for this phenomenon, need further studies. In contrast, results in the GTT group suggests that supplementation with γ-tocotrienol alone was not inhibitory and may even aid in normalizing the vitamin D metabolism. Thus, the inhibitory effect of the TOC supplementation was probably due to its a-tocopherol component. Urine calcium was found to be decreased in the VED and ATF groups post-treatment compared to baseline. Because vitamin E deficiency inhibited the activation of vitamin D,[18] and 1,25-dihydroxyvitamin D3 is needed for optimum intestinal calcium absorption, these data suggests that hypocalcemia was induced in all the vitamin E-deficient groups. However, the group supplemented with GTT was able to correct its hypocalcemia by the second month, thereby resulting in the normal serum and urine calcium levels. This was probably because supplementation with γ-tocotrienol allowed for the activation of vitamin D. However, for the VED and ATF groups, urine calcium was low, suggesting that supplementation with a-tocopherol alone was not effective in normalizing calcium homeostasis. Probably an increased calcium reabsorption by the kidneys occurred in compensation. For the ATF group, the low levels of serum 1,25-dihydroxyvitamin D3 made this problem worse. In the TOC group, a combination of the effects of γ-tocotrienol and a-tocopherol were seen, that is, serum 1,25-dihydroxyvitamin D3 was low, but urinary calcium concentration was normal. Tocotrienols are less excreted compared to tocopherols[20] and its protective effects against vitamin E deficiency may be prolonged. Others have found that vitamin E deficiency reduced the a-tocopherol binding protein in liver.[21] This may explain the lower efficacy of a-tocopherol compared to palm tocotrienol. Tocomin ® supplementation caused a reduction in 1,25-dihydroxyvitamin D3 level as in the ATF group and showed comparable effect on urine level of calcium as in GTT group. Tocomin ® contains 22.1% γ-tocotrienol and 11.5% a-tocopherol. This composition of Tocomin ® may have caused the mixed effects seen in the serum 1,25-dihydroxyvitamin D3 level and urine calcium level. This implies that supplementation of GTT alone is superior to that of the mixture of a-tocopherol and γ-tocotrienol. Furthermore, the quantity of GTT present in Tocomin ® is lower than the pure γ-tocotrienol thus, diminishing its effects. Furthermore, the presence of a-tocopherol in TOC may contribute to its lower efficacy. Therefore, the overall picture seen with TOC supplementation will depend on the dominant isomer, as well as on the duration of treatment. Our findings also suggest that a-tocopherol and γ-tocotrienol might act via different pathways to influence vitamin D metabolism and calcium homeostasis. Another possible explanation is that the tissue distribution of these forms of vitamin E varies between each other and exerts its action on different tissues depending on their distribution. One study showed that the distribution of tocotrienol and tocopherol is tissue specific.[22] In the present study, the serum parameters were measured at the beginning and at the end of the treatment period. Any differences in the parameters levels, which may occur between these two time-points, might be overlooked. It is highly recommended that a time-course measurement be done in future studies. Vitamin E deficiency (VED group) impaired calcification of both femur and fifth lumbar vertebra bones as compared to BC suggesting an impaired calcification or calcium loss. However, the lumbar bone calcium content of the VED group, but not femur, differed significantly compared to RC. This is in agreement with our previous study and the bone loss observed may be due to bone resorption induced by secondary hyperparathyroidism.[19] Furthermore, inhibition of vitamin D metabolism is known to impair calcium absorption from the intestine; therefore less calcium was available for deposition into bone. Lumbar vertebral bones consist mainly of cancellous bone, while femoral bones contain more cortical bone. Cancellous bone has a higher metabolic and remodeling rate compared to cortical bone, and therefore, the lumbar vertebral bones are more affected by any stress or changes compared to femoral bones.[23] Cancellous bone has a higher ratio of bone surface area to bone volume compared to cortical bone. Bone remodeling occurs on the bone surface; hence cancellous bone metabolic rate is tenfold higher than cortical bone.[24] As a result, the effects of vitamin E deficiency on bone calcium content were more apparent in the lumbar vertebral bones as compared to the femoral bones. The RC group showed an increase in the femur and lumbar bone calcium content as compared to the BC group indicating that the rats were growing. Bone calcium content of the VED group did not increase compared to BC and the lumbar bone calcium content was lower than RC. This suggests that vitamin E deficiency impaired bone calcification. Femur bone calcium content in the vitamin E supplemented groups did not differ compared to VED. In the lumbar bones, GTT and TOC group failed to increase its bone calcium content to the level of RC although GTT group was able to maintain biochemical parameters comparable to the RC group. The short treatment period may not be sufficient for GTT to reverse the effects of vitamin E deficiency on bone calcium content. Previous studies showed that palm vitamin E supplementation prevented bone calcium loss in rats exposed to ferric-nitrilotriacetate, an oxidizing agent.[8] Palm oil has high quantity of tocotrienol and any protective effects shown by palm vitamin E is probably due to its tocotrienol component. Another study showed that palm vitamin E supplementation, but not a-tocopherol, at the dose of 30 and 60 mg/kg for 8 months was able to increase bone calcium content in vitamin E-deficient rats.[14] The beneficial effects of GTT on bone calcium in this study could have been more obvious had the study period been longer. ATF supplementation seemed to improve lumbar bone calcium content, but this group failed to normalize the levels of biochemical parameters (serum 1,25-dihydroxyvitamin D3 and urine calcium) even after 2 months. The long-term impact needs to be elucidated in further studies. Longitudinal growth of the femur in all of the groups was increased compared to the BC group. The differences in femur length observed in this study were almost the same as the femur bone calcium content results. No difference was seen between the VED group and the vitamin E supplemented groups. However, a study done in chicks found that vitamin E improved bone growth by increasing the thickness of the growth plate cartilage.[25] Conclusion Our data indicates that vitamin E deficiency inhibits activation of vitamin D, which leads to hypocalcaemia and bone loss. Supplementation with γ-tocotrienol 60 mg/kg rat weight, but not a-tocopherol 60 mg/kg rat weight or a mixture of a-tocopherol and the tocotrienols as in Tocomin ®, preserved vitamin D metabolism and maintained calcium homeostasis. Therefore, gammα-tocotrienol and not alpha-tocopherol protects female rats from the effects of vitamin E deficiency on vitamin D metabolism; however, it does not prevent a decline in lumbar vertebra calcium content over a 2-month period.Acknowledgments The authors would like to thank Universiti Kebangsaan Malaysia for the research grant FF-012-2002. We also thank carotech Sdn Bhd for supplying gammα-tocotrienol and Tocomin ® needed for the study. We express our gratitude to Mr. Mohd Imran Mohd Ali and Pn Azizah Osman for their technical assistance.References
Copyright 2005 - Indian Journal of Pharmacology The following images related to this document are available:Photo images[ph05080t2.jpg] [ph05080t1.jpg] [ph05080t3.jpg] [ph05080t4.jpg] |
| |||||||||

![[Table - 1]](/showimage?ph/photo/ph05080t1.jpg){kind=link}
![[Table - 2]](/showimage?ph/photo/ph05080t2.jpg){kind=link}
![[Table - 3]](/showimage?ph/photo/ph05080t3.jpg){kind=link}
{kind=link}