 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Indian Journal of Pharmacology, Vol. 37, No. 6, November-December, 2005, pp. 366-370 Research Paper Effect of reactive oxygen species on cholinergic receptor function Venkatesham A, Babu PSharath, Sagar JVidya, Krishna DR Cancer, Aging and Metabolism Research Division, University
College of Pharmaceutical Sciences, Kakatiya University, Warangal -506 009,Andhra
Pradesh, India Code Number: ph05097 Abstract OBJECTIVE: To investigate the role of reactive oxygen species (ROS) on cholinergic receptor function.MATERIALS AND METHODS : Rectus abdominis and isolated heart preparations of frog ( Rana tigirina ) were used to assess nicotinic and muscarinic receptor activity, respectively. Thirty percent hydrogen peroxide (H2O2) solution and Fenton mixture (Fm, 13.9 mg, 50 µM of FeSO4, 75 mg of sodium EDTA and 50 µL of 30% H2O2 were added to 10 ml of 0.1 M K2HPO4) were used to generate 1 mM H2O2 and hydroxyl free radicals. The responses were recorded with acetylcholine at different phases of exposure of tissues to ROS. Normal frog Ringer was used as a physiological solution. Responses of acetylcholine were also recorded in the presence of ROS before and after exposure of the tissue to an antioxidant (ascorbic acid). RESULTS : Free-radical-mediated receptor damage was dose (1-100 mM H2O2) and time (10-30 min) dependent when responses were taken with 30 µg and 30 ng of ACh for nicotinic and muscarinic receptors, respectively. There was no effect of ROS on prior exposure of tissue to ascorbic acid (antioxidant) at a concentration of 300 µg/ml. The antioxidant has not shown any beneficial effect on sulfhydryl groups of G-protein-coupled muscarinic receptors, which are more susceptible and sensitive to ROS than ion-channel nicotinic receptors where there is 96% protection with the antioxidant. Reactive oxygen species has shown different effects on receptor function. CONCLUSION: Free radicals continuously cause considerable damage to the receptors. G-protein-coupled muscarinic receptors are more susceptible than ion-channel-linked nicotinic receptors. Antioxidants are shown to play a major role in protecting free-radical-mediated receptor damage. Keywords: Antioxidants; free radical; acetylcholine; ascorbic acid. Introduction Receptors are specialized to recognize and respond to individual signaling molecules with great selectivity. Proteins, glycoprotein, proteolipids, and associated proteinaceous matter constitute receptors.[1] Free radicals are chemical species possessing an unpaired electron that can be considered as a fragment of molecules which are generally very reactive. They are produced continuously in cells either as accidental by-product of metabolism or deliberately during phagocytosis. Reactive oxygen species (ROS) includes not only oxygen free radicals (OFR) but also nonradical oxygen derivatives that are involved in oxygen radical production. Hydrogen peroxide (H2O2) easily breaks down, particularly in the presence of transition metal ions to produce the most reactive OFR, the hydroxyl radicals. Oxygen-derived free radicals and other reactive oxygen species have been shown to be important mediators of cellular and tissue injuries in a variety of diseases such as diabetes, heart failure, hypertension, and ischemia/reperfusion. Further, ROS are mediators of lipid peroxidation, protein and nucleic acid modification, which can result in altered cellular responses and cell death.[2] It has been recognised that free radicals are involved in the etiology of many diseases. Although insight into the role of free radicals in physiology and pathogenesis has been gained over the recent years, remarkably little knowledge exists on the effect of free radicals on receptor-mediated response.[3] Imbalance between production of OFR and antioxidant defense can result in oxidative stress leading to metabolic impairment and cell death. Oxidative stress may be due to deficiency of antioxidants (such as glutathione, ascorbate or a-tocopherol), antioxidant enzymes [superoxide dismutase (SOD), catalase, glutathione peroxidase] and/or from increased levels of OFR.[4] Several definable diseases arise from disorders in receptors or receptor-effector systems, such as feminization syndrome, generalized endocrinopathy and pseudohypo-parathyrodisim type 1a.[5] Nicotinic receptors in myasthenia gravis,[6] LDL receptors in familial hypercholesterolemia, V2 receptors in nephrogenic diabetes insipidus, ACTH receptors in cortical insufficiency, PTH receptors in pseudohypo-parathyroidism and β-adrenoceptors in hypertensive states are implicated in the free radical damage of receptors.[7] The generation of ROS has been observed under various pathological conditions. In rat atria exposed to ROS, adrenoreceptor stimulation reduces the contractile force as a result of a protein kinase C-mediated Na+/K+-ATPase activation.[8] Activation of the M1 muscarinic receptor subtype in rat pheochromocytoma cells stably expressing cloned M1 muscarinic acetylcholine receptors was previously shown to induce morphological changes and growth arrest. However, signalling pathways, which led to these effects, were not identified.[9] So far, the influence of ROS on the cardiac muscarinic receptors has not been studied in detail. The only studies so far performed indicate that the free radicals and H2O2 can alter the binding characteristics of the cardiac M2 receptor, while the functional data are not yet available.[10] Reactive oxygen species act as second messengers in muscarinic-induced cellular signalling. Moreover, generation of ROS appears to be an early and critically intermediary event, which occurs immediately after stimulation of the muscarinic receptor and in turn affects the muscarinic-mediated cellular signalling.[9] Thus, receptors are prone to be damaged by the effects of free radicals. Even the pathogenesis of free-radical-mediated receptor dysfunction on cholinergic receptors is not well defined. In the present study, an attempt has been made to study these effects in vitro . Materials and methods Drugs and their source Preparation of solutions Frog Ringer solution : The physiological salt solution was prepared by adding NaCl 110 mM, KCl 1.9 mM, CaCl2 1.1 mM, NaHCO3 2.4 mM, and Glucose 11.1 mM in distilled water. Acetylcholine solution : The stock solution of acetylcholine was prepared in 5% NaHPO4 solution so as to contain 10 mg/ml and it was stored at -4°C. Serial dilutions of 1 mg/ml, 100 mg/ml, 10 mg/ml, 1 mg/ml, and 100 ng/ml were prepared from the stock solution at the time of the experiment.[12]
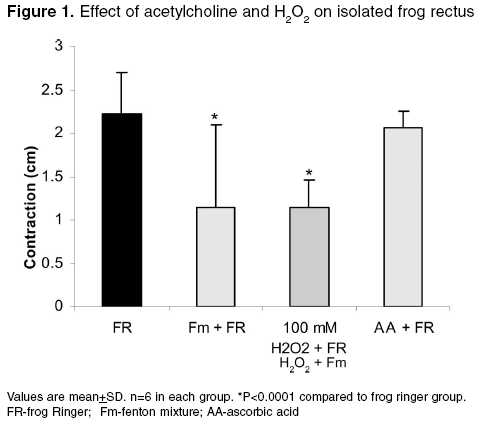
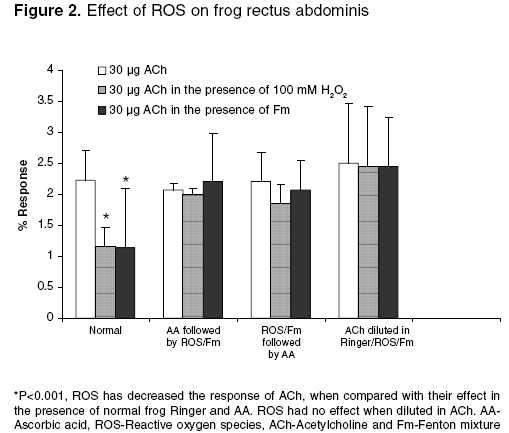
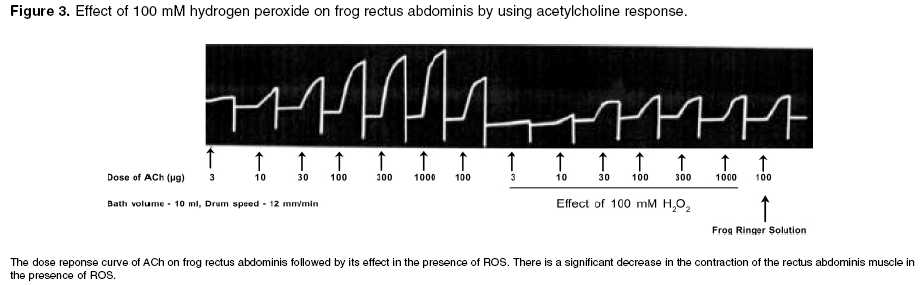
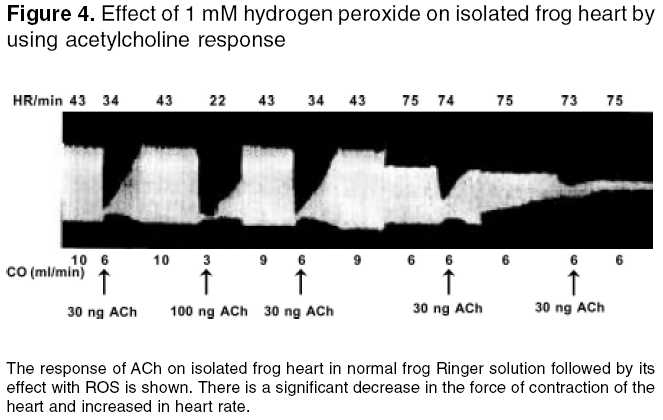
Effect of free radicals on nicotinic and muscarinic receptor function Effect of pretreatment with AA (antioxidant) on ACh in the presence of 100 mM H2O2 and Fm on FRA: The dose response curve was taken in the presence of frog Ringer solution containing AA (300 µg/ml) after remaining in contact with the muscle for 10 min. After that, the frog Ringer solution containing AA was replaced with Ringer solution containing 100 mM H2O2 or Fm. The rectus abdominis muscle was bathed in the above-mentioned solution for 10 min. Then, the responses were recorded. Effect of ACh in the presence of 100 mM H2O2 followed by treatment with AA on FRA: The dose response curve was recorded in normal frog Ringer solution. After that, the frog Ringer solution was replaced with frog Ringer solution containing 100 mM H2O2 and the rectus abdominis muscle was bathed in the above-mentioned solution for 10 min and the responses were recorded. Then, the above solution was replaced with frog Ringer solution containing AA and responses were recorded. Effect of ACh diluted in 100 mM H2O2 and Fm on FRA: The dose response curve was recorded on FRA with ACh diluted in frog Ringer solution, 100 mM H2O2 and Fm separately and compared with dilutions of ACh in frog Ringer solution. Effect of free radicals on muscarinic receptor function Effect of pretreatment of AA on ACh in the presence of 1 mM H2O2 and Fm separately on isolated frog′s heart: ACh response was recorded in the presence of frog Ringer solution containing AA (300 µg/ml) after stabilization of the heart for 10 min. After that, the frog Ringer solution containing AA was replaced with frog Ringer solution containing 1 mM H2O2 and Fm separately. After stabilization for 10 min, ACh responses were recorded on the kymograph. Effect of ACh in the presence of 1 mM H2O2 followed by treatment with AA on isolated frog heart: ACh response was recorded in frog Ringer solution after stabilization of heart for 10 min. After that, the responses were recorded with frog Ringer solution containing 1 mM H2O2. Then, the above solution was replaced with frog Ringer containing AA and after stabilization for 10 min the ACh responses were recorded again. Effect of ACh diluted in 1 mM H2O2 and Fm separately on isolated frog heart: 10, 30, 100, and 300 ng of ACh were prepared in frog Ringer solution, 1 mM H2O2 and Fm responses were recorded separately on isolated frog′s heart and compared with dilutions of ACh in frog Ringer solution. The study was carried out after clearance by the institutional animal ethics committee. Statistical analysis Results Effect of 100 mM H2O2 and Fm on nicotinic receptor
function Effect of 1 mM H2O2 and Fm separately on muscarinic receptor function Discussion It is clearly evident that free radicals play a pivotal role in the etiology of many diseases. Local free radical formation that occurs under certain pathological conditions, like ischemic reflow, may regulate receptor response.[3] Free radical susceptibility of receptors can be deduced from the presence of functionally critical sulfhydryl group located in the receptor proteins.[15] In the present study, it has been observed that the free radical generation and loss in receptor function were concentration-dependent, which has earlier been reported by Olszewski et al .[16] on trachealis muscle of horse, suggesting that more generation of the free radicals leads to greater receptor damage, which may ultimately result in reduced receptor function. Further, the concentration-dependent effect of ROS was also observed in both the nicotinic receptors of FRA and muscarinic receptors of isolated frog heart, which is thought to be due to the involvement of sulfhydryl groups; it has been proposed for muscarinic cholinergic receptors and detailed molecular delineation of the cysteine residues involved in disulfide bonding of the receptor protein was studied.[15] Ashkenazi et al . reported that ROS potentiate the negative inotropic and attenuate the positive inotropic signalling events in M2 receptor, thereby potentiating the negative inotropic effect of muscarinic receptor agonist (ACh) in isolated rat left atria. These actions of the M2 receptor are thought to be mediated by inducing phospholipase C-mediated phospoinositide turnover.[17] Doleman et al ., in 1988 reported the antioxidant effect of vitamin E and selenium in lung tissue after exposure to ROS[18] and Mangelus et al ., in 2001 also reported that the Ras, extra cellular signal-regulated kinase and p38 are responsible for the imbalance which is brought about by muscarinic activation. These pathways were blocked by the antioxidant N-acetylcysteine when exposed to ROS.[9] In the present study on pretreatment of frog′s rectus abdominis with AA, there was no reduction in ACh response with frog Ringer containing ROS. It is further evident that there is no effect of ROS on nicotinic receptor function on prior exposure to antioxidants (AA). In contrast, AA has not shown any beneficial effect on the muscarinic receptor site. The study on exposure of antioxidants after treatment with ROS showed that the receptor activity has been reversed in the nicotinic receptor in contrast to the muscarinic receptor, which may be due to permanent loss of muscarinic receptor activity. This again implies that the muscarinic receptors are more susceptible to free radicals than the nicotinic receptors. It may be attributed to the sulfhydryl groups of the G-protein-coupled muscarinic receptors, which are more susceptible and sensitive to ROS than the ion-channel-linked nicotinic receptors. The investigation also reveals that, with increase in exposure time (10-30 min) of ROS at the receptor site, there is a corresponding decrease in receptor activity even at a higher dose of ACh (300 µg). This clearly indicates that effective free radical attack is time dependent. The time-dependent effect of ROS was observed both in nicotinic and muscarinic receptors. The effect of ACh was reduced with very low concentration of ROS (1 mM H2O2, 1:50 Fm) on muscarinic receptors, compared to that on nicotinic receptors (0.1 M H2O2 or 3:50 Fm). This also indicates that the muscarinic receptors are more sensitive to ROS than nicotinic receptors. Reactive oxygen species has shown different effects on receptor function. The actions of Fm were reversed both in nicotinic and muscarinic receptors when replacing with normal frog Ringer′s solution but not with H2O2. This indicates that H2O2 causes permanent damage at the receptor site. ACh diluted in Fm and H2O2 showed no change in ACh response, which indicates that the effect of ROS was due to altered receptor function but not due to chemical alteration of ACh or in vitro interaction. Conclusion From the above study, it is clear that free radicals cause damage to the receptors. G-protein-coupled muscarinic receptors are more susceptible than ion-channel-linked nicotinic receptors. Ascorbic acid, a free radical scavenger, offers protection from ROS at the receptor site. A thorough and detailed investigation had been planned to support the study.Acknowledgments The first author is thankful to the University Grants Commission (UGC), New Delhi, for providing financial assistance in carrying out the work.References
Copyright 2005 - Indian Journal of Pharmacology The following images related to this document are available:Photo images[ph05097f2.jpg] [ph05097f1.jpg] [ph05097t1.jpg] [ph05097f4.jpg] [ph05097f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}