 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
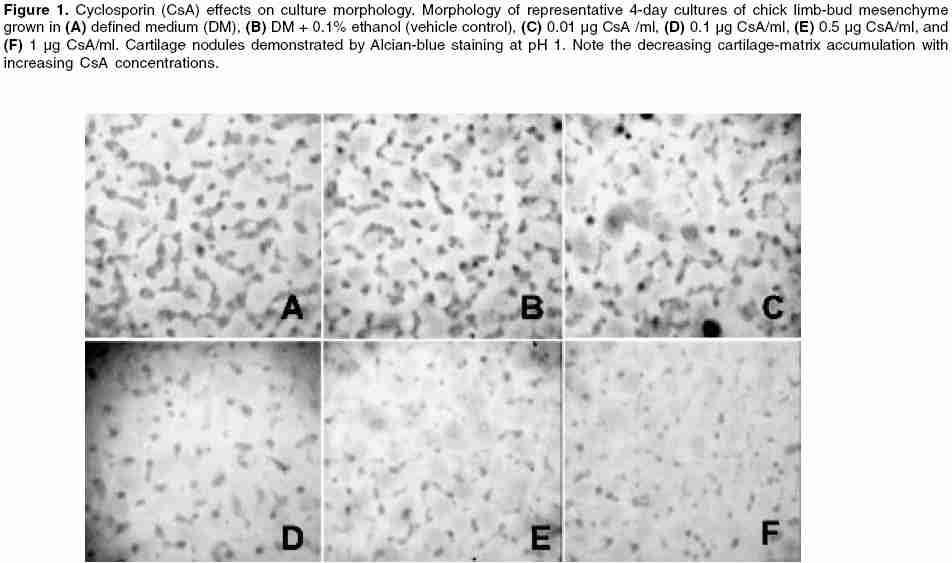
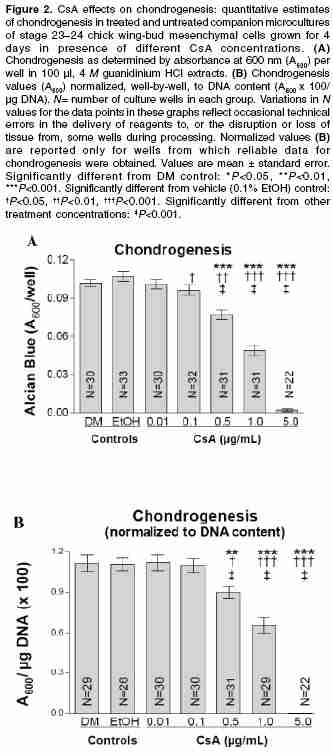
Indian Journal of Pharmacology, Vol. 38, No. 1, January-February, 2006, pp. 43-48 Research Paper Inhibition of chondrogenic differentiation in chick limb-bud mesenchyme microcultures treated with cyclosporine Bahmanpour1 Soghra, Paulsen2 Douglas F 1Anatomy Department, Shiraz University of Medical Sciences, Shiraz, Iran. 2Departments of Anatomy and Neurobiology, Moorehouse School of Medicine, Atlanta, GA, USA. Code Number: ph06008 Abstract Objectives: To explore the effects of cyclosporine (CsA) on skeletal development (chondrogenesis).Materials and Methods: Mesenchymal cells obtained from stage-23 to stage-24 chick-embryo limb buds were grown in 96-well plates using chemically defined tissue-culture medium. Cultures were treated with CsA (0.01-5.0 µg/ml) and incubated (37ºC, 5% CO2) with daily medium changes for 4 days. After incubation of the cells in multiwell plate, cartilage differentiation (chondrogenesis) was assessed by selectively staining sulfated glycosaminoglycans (GAGs) in the cartilage matrix with Alcian blue, extracting the GAGs with 4 M guanidinium HCl, and spectrophotometric analysis of the extracts. Results: CsA treatment had concentration-dependent effects on chick limb-bud mesenchymal cell cultures. At 5 µg/ml, CsA caused cell loss, as judged microscopically by the paucity of cells remaining at the end of the culture period. CsA concentrations between 0.1 and 1 µg/ml caused a marked, dose-dependent decrease in chondrogenesis. At 0.01 µg/ml, CsA had no significant effect on chondrogenesis. At concentrations above 0.01 µg/ml, normalized data showed significant chondrogenic inhibition at 0.5 and 1.0 µg/ml CsA. Conclusions: The findings suggest a possible biological basis for CsA-associated effects on mesenchyme-derived tissues and provide a model system for further studies. Keywords: Chondrogenesis, skeletal development spectrophotometric analysis Introduction Cyclosporine-A (CsA) therapy is a common and effective immunosuppressive treatment for transplant recipients and, increasingly, for autoimmune disorders.[1] Recently, CsA has also been shown to reduce CD4+ T-cell loss in HIV-infected patients.[2] CsA inhibits T-lymphocyte activation by inhibiting calcineurin activity, and thereby inhibiting cytokine production (e.g., interleukin-2 and interferon-γ).[3],[4] Calcineurin is a heterodimeric, calcium-dependent serine/threonine phosphatase, whose wide distribution in vertebrate tissues may account for CsA′s effects on many nonlymphoid cell functions.[5],[6] Indeed, CsA is often used as a specific inhibitor in studies of calcineurin′s regulatory effects in a variety of tissue types. [6],[7],[8] CsA′s increasing use in reproductive-age women and its apparent capacity for effects on a variety of tissues have raised concerns about its teratogenic potential. Reports of major malformations in live human births after CsA exposure during pregnancy are rare. Yet, understandable limitations on randomized, controlled studies in humans; the absence of comparisons of exposure during embryonic and fetal periods, confounding influences of other drug therapies that accompany CsA treatment (e.g., glucocorticoids). Variable CsA dosage and pharmacokinetic patterns, and trends toward increased risk of prematurity, low birth weight, and congenital malformation-all indicate a continuing need both for caution in prescribing CsA and monitoring during pregnancy and for further studies on the effects of CsA on embryonic and fetal tissues.[9] Although there is evidence that placental transfer of CsA is limited in rabbits and rodents,[10] its ability to reach human embryos and fetuses has been established.[11] At least one case of CsA-induced osseous malformations, including limb defects, has been reported.[12] When vertebrate embryos have been directly exposed to CsA, significant anomalies in the heart and other mesenchyme-derived tissues and organs have occurred.[6],[8],[11],[13] Adult skeletal and connective tissues exposed to CsA show alterations in extracellular matrix synthesis and secretion,[14],[15] suggesting a possible mechanism for altered development when calcineurin signaling is disrupted. Recently, calcineurin mRNA expression has been demonstrated in chick limb-bud mesenchyme.[8] Thus, analyses of CsA effects on extracellular matrix synthesis and secretion in cultures of limb-bud mesenchyme may improve our understanding of CsA′s capacity to disrupt normal development. A further aim of this study was to adapt and validate a modified microtiter micromass culture model[16] to explore CsA effects on in vitro cartilage differentiation (chondrogenesis). Micromass cultures of embryonic limb-bud mesenchyme have been used and adapted to study many factors affecting differentiation.[17] Mesenchymal cells isolated from embryonic limb buds undergo differentiation in tissue culture following a sequence of steps remarkably similar to those occurring in vivo .[17],[18] In part because of their accessibility, chick limb-bud mesenchymal cells have been especially useful in studies of differentiation mechanisms and in screening drugs and other compounds for toxicity and teratogenicity. [17],[18],[19] Extracellular accumulation of cartilage proteoglycan aggregates (aggrecan) in these cultures serves as the primary endpoint for assessing chondrogenesis.[15],[16],[17],[18] Cartilage proteoglycans consist primarily of chondroitin-sulfate glycosaminoglycan (GAG) side chains covalently linked to a core protein. Individual proteoglycans interact with hyaluronan to form large proteoglycan aggregates. Proteoglycans of noncartilage connective tissues form smaller aggregates and have a different composition with respect to types and relative amounts of GAGs and core proteins.[20] The amount of aggrecan accumulated in the extracellular matrix can be assessed by staining the culture with Alcian blue at pH 1, limiting its binding to sulfate groups in the matrix GAGs.[16] Compounds that interfere with cartilage differentiation affect aggrecan accumulation, and thus the amount of Alcian-blue stain specifically bound by the cultures. Changes in Alcian-blue staining in CsA-treated cultures therefore reflect effects on cartilage differentiation from mesenchyme and may provide insight into CsA′s potential for disrupting skeletogenesis. Materials and Methods Tissue culture The wing buds were collected using a Pasteur pipet, delivered to a 15 ml centrifuge tube (Corning) containing 5 ml room temperature TBSS, and allowed to sediment at 1 g. The supernatant was removed and discarded (further reducing blood cell contamination) and the limb-buds were resuspended in 2 ml of trypsin-collagenase (0.1% trypsin [GIBCO/BRL] and 0.1% collagenase [Worthington], with 2% heat-inactivated chicken serum in calcium- and magnesium-free TBSS). The limb buds and enzyme solution were then delivered to a sterile 25 ml Erlenmeyer flask and rotated at 70 rpm in a 37°C water bath. After 3 min, the limbs were triturated with a Pasteur pipette to obtain a single-cell suspension. Occasionally, the dissociation was visually judged incomplete and the flask and its contents were returned to the water bath for an additional 1-3 min. After a final trituration, the suspension was passed through a 20 µm nylon mesh filter (Nitex) to remove any remaining cell aggregates and sheets of ectoderm; 5 ml of unsupplemented F12/DMEM (50 : 50, Cellgro) were added to the suspension to dilute the enzyme. The resulting single-cell suspension was pelleted and resuspended in 5 ml of complete defined medium (DM, F12/DMEM supplemented with 100 n M hydrocortisone (Sigma), 5 µg/l insulin (Collaborative Research, MA), 5 µg/ml chicken transferrin (conalbumin, Sigma), 1 mg/ml l-ascorbic acid (Sigma), and the antibiotics penicillin, streptomycin, and fungizone (GIBCO)[16] to further dilute the enzyme and facilitate cell counting. The cell density of the resulting suspension was determined using a hemacytometer. Trypan-blue exclusion testing showed that this dissociation procedure yields suspensions with more than 95% viability. After counting, the cells were pelleted and resuspended in an appropriate amount of DM to yield a density of 1 x 106 cells/ml. Aliquots (250 µl) of this suspension (2.5 x 105 cells) were inoculated into each well of a 96-well, tissue-culture-treated, flat-bottomed, microtiter plate (Costar). This plating procedure represents a departure from previous studies in which a 10 µl droplet containing the same number of cells was delivered to each well.[16] The modification was made for several reasons. It is difficult to center the droplet in these wells. If the droplet touches the wall, its surface tension is disrupted, resulting in a crescent of cells along the contacted wall. In addition, because there is no room in the small wells to add medium to counteract evaporation during the attachment period, there were concerns that evaporation from such small drops would increase the concentration of the many medium components during attachment, producing effects that might be inconsistent from well to well and difficult to assess. Delivering 250 µl aliquots to each well results in a more homogeneous and reproducible cell distribution over the well surface. Obviously, the plating density is reduced by this modification. However, as the data reported demonstrate, this modified procedure results in measurable chondrogenesis and DNA content. Further, because it was unknown prior to this study of whether CsA would enhance or inhibit chondrogenesis in these cultures, or whether there would be an effect on cell proliferation, the lower density provides the added advantage of assessing changes in either direction. After seeding, the plates were incubated at 37°C in a water-jacketed incubator containing a humidified atmosphere of 5% CO2 for a period of 2 h to allow initial attachment before adding the treatments. CsA treatment. Because CsA may cause hepatotoxicity and nephrotoxicity in patients undergoing immunosuppressive therapy, monitoring of circulating CsA concentrations is often requested in order to maintain a therapeutic range.[5] However, individual differences in pharmacokinetics[5] and concerns about the accuracy of the methods used to monitor CsA concentration in the blood[7] makes it difficult to pinpoint ideal therapeutic doses or circulating concentrations. Nevertheless, circulating CsA concentrations have been reported to range between 0.1 and 2.0 µg/ml, although substantially higher concentrations have been detected in some individuals.[5],[22] The CsA concentrations used for this study were chosen to bracket this range. CsA was obtained as a purified powder (Sigma, St. Louis, Missouri). Fresh treatments were prepared for each experiment because preliminary studies showed a diminished effect in cultures treated with stocks older than 1 week (data not shown). A primary stock solution of 5 mg/ml CsA in 100% ethanol (EtOH) was prepared. A secondary stock containing 50 µg/ml CsA was then prepared by dilution in DM. Additional stocks were prepared by dilution in DM to create treatments containing the various CsA concentrations (0.01, 0.1, 0.5, 1, and 5 µg/ml). The vehicle control treatment was prepared containing EtOH equivalent to that in the 5 µg CsA/ml treatment (0.1% EtOH in DM). After the attachment period, 25 µl of a 10X concentration of the appropriate treatment was delivered to the plating medium (DM) in each well. The cultures were grown for 4 days with careful daily complete replacement with 250 µl of DM alone or containing the appropriate treatment, or vehicle control medium. Chondrogenesis measurements Statistical methods Results Controls Chondrogenesis Because cell density is an important factor during chondrogenesis in limb-bud mesenchymal cell cultures, normalizing quantitative measures of chondrogenesis to DNA content (our previous data) can be helpful in assessing the extent to which the observed effects may be attributable to alterations in cell density. The method described allows chondrogenesis measurements to be obtained for each well rather than companion cultures, providing an internal control for the effects of cell density. Values were normalized for each well by dividing the net A600 for the extracted Alcian-blue stain by the µg DNA (previous data) and multiplying the result by 100. This creates a separate and perhaps more reliable data set for evaluating treatment effects on chondrogenesis. These normalized values showed significant inhibition of chondrogenesis for CsA treatments at 0.5, 1.0, and 5.0 µg/ml, as compared with either the DM-only control ( P< 0.005, P< 0.001, P< 0.001, respectively) or the vehicle control [ P< 0.05, P< 0.001, P< 0.001, respectively; [Figure - 2]B. Spearman rank correlation testing again confirmed that CsA-mediated inhibition of chondrogenesis to be concentration-dependent [ vs DM-only and vs vehicle control: P = 0.005, r = -0.943; [Figure - 2]B. Effects at 5.0 µg CsA/ml, although significant, were viewed as uninterpretable because of the paucity of cells remaining at the end of the culture period. Discussion The effects of CsA demonstrated in the present study on in vitro chondrogenesis shows a concentration-dependent inhibition and supports the view that immunosuppressive agents affect cell differentiation in cells and tissues other than their intended lymphoid targets.[5],[6] Despite the consensus that CsA affects cell function through interactions with calcineurin,[4] the outcome of CsA treatment at the cellular level is highly variable. Apparent inconsistencies in the reported effects likely reflect tissue- and cell-type-specific responses to CsA treatment, confirming the importance of individual studies of potential target systems. In studies of adult and embryonic skeletal and connective tissues, CsA has been reported to both enhance and inhibit extracellular matrix (ECM) production. CsA-mediated enhancement of ECM production by fibroblasts has been demonstrated for renal and gingival tissues.[13],[14] Several reports provide evidence of CsA-mediated disruption of the balance between ECM synthesis and degradation in bone. In rodents, CsA treatment causes severe osteopenia in sites of high bone turnover. In vitro studies on osteoblast cell lines have shown that CsA alters ECM synthesis and degradation. It inhibits osteoblast proliferation, alkaline phosphatase activity, and cell attachment without affecting parathyroid hormone binding or cAMP levels.[25] In another study, CsA was also shown to decrease collagen type 1 production and to increase matrix metalloproteinase activity.[26] On the other hand, CsA inhibits chicken osteoclast differentiation and bone resorption in vitro .[27] All these studies are in accordance with the present study, which confirms the inhibitory effects of CsA on chondrogenesis. Indeed, the CsA decreased the ECM productin and cell differentiation. Reports of CsA′s effects on cartilage and chondrogenesis are rare. A case report of skeletal malformations in an infant born to a woman on CsA therapy[11] is suggestive of an effect on chondrogenesis but is hardly conclusive and does not address etiology. CsA inhibits mandibular condylar cartilage maturation in young rats,[28] but actually enhances chondrogenesis and osteogenesis during repair of surgically created mandibular defects in older animals receiving human demineralized bone grafts;[29] an effect suggested to reflect CsA-mediated suppression of the immune response to the xenograft. Miniero et al. published a retrospective survey in Italy reporting that 29.2% of pregnant women that had taken immunosuppressive experienced miscarriage.[30] Miscariage could be due to congenital defects leading to termination of pregnancy. In another study, Miniero found out that out of eight infants born from transplant recipient mother, one had congenital talipes valgus.[31] His finding confirms the CsA effects on congenital abnormalities and skeletal malformation and it is in accordance with the present study. CsA′s ability to disrupt embryonic chondrogenesis, as indicated by effects on nodule morphology and inhibition of matrix accumulation, seems clear under the present experimental conditions and previous study, which showed the CsA effects on cell proliferation and therefore cells and ECM production were reduced under CsA therapy.[21] However, chondrogenesis is a complex process involving the spatiotemporal coordination of many cellular and molecular mechanisms.[17] The observed disruption of cartilage histogenesis and cytodifferentiation may reflect direct effects on gene expression and/or more indirect effects on cell adhesion, cellular rearrangements, and interactions required for nodule formation, cellular metabolism, or aspects of matrix synthesis, secretion, assembly, or turnover. An obvious starting point for studies of the mechanism underlying CsA-mediated disruption of chondrogenesis is the calcineurin signaling pathway. In mouse limb-bud micromass cultures, calcineurin overexpression has been reported to enhance chondrogenesis through interactions with the nuclear factor of activated T-cells (NFAT) isoform, NFAT4 (x, c3), and downstream effects of the activated transcription factor complex on BMP2 production.[32] The specificity of this signaling mechanism is underlined by evidence that NFAT1 (p, c2) actually inhibits chondrogenesis.[33] Upstream activators of the calcineurin/NFAT signaling pathway, such as the Wnt family of signaling molecules,[7] are known to have important effects on chondrogenesis and cellular adhesion.[34] Misexpression of Wnt-7a, but not Wnt-5a, strongly inhibits the chondrogenesis of primary chick limb mesenchymal cells in vitro .35 Additional studies of calcineurin and NFAT-isoform expression patterns, their transcriptional targets, as well as CsA′s role in modulating calcineurin/NFAT-isoform association at different stages of skeletal development will improve our understanding of chondrogenesis and may provide further guidance in the use of CsA as a therapeutic agent. Conclusion CsA′s effects on chondrogenesis in this study provide a further example of its effects on nonlymphoid tissues. CsA caused a concentration-dependent inhibition of chondrogenesis in 4-day cultures of chick limb-bud mesenchymal cells isolated from stage-23 to stage-24 chick embryos. Its most significant inhibitory effects occurred at treatment concentrations of 0.5 and 1.0 µg/ml, which fall within the expected circulating levels in patients receiving immunosuppressive CsA therapy. Whether this inhibition of chondrogenesis reflects an influence on cell adhesion, the cellular rearrangements necessary to form the cartilage nodules, cellular metabolism, matrix synthesis and secretion, or more direct effects on gene expression remains to be determined. Nevertheless, the findings provide new information regarding CsA′s effects on chondrogenesis from embryonic mesenchyme and appropriate CsA concentrations for use in further studies. In addition, they validate a useful model system for pursuing studies of CsA′s effects on these mechanisms.Acknowledgments This study was supported by NIH grants GM08248 and RR03036 to the Morehouse School of Medicine (MSM) and by sabbatical support to SB from Shiraz University of Medical Sciences (SUMS). The authors thank Dr. Mary Scanlon and Andrew Shaw in MSM′s Image Analysis Facility for advice and assistance, and Dr. N. Zare, F. Dehbozorgian and A. Baghaei in the SUMS Statistical Consultation Center for assistance with the statistical analysis. We also thank Dr. Comnuan Nokkaew and Candace Marshall for technical assistance.References
Copyright 2006 - Indian Journal of Pharmacology The following images related to this document are available:Photo images[ph06008f2.jpg] [ph06008f1.jpg] |
| |||||||||

{kind=link}
{kind=link}