 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
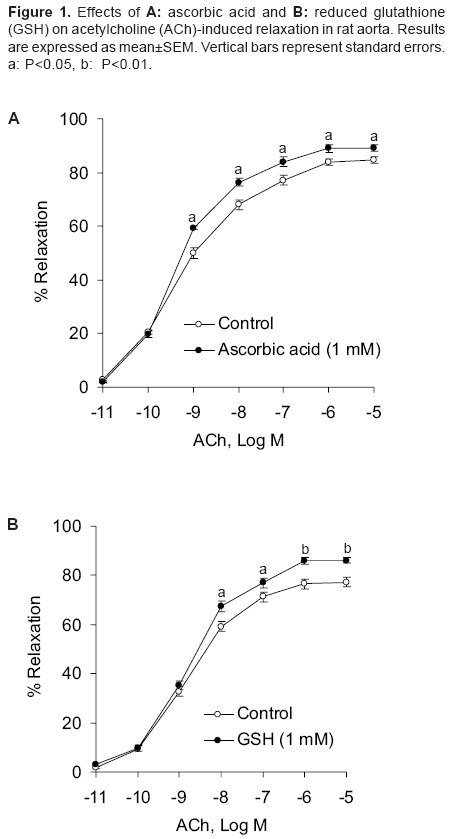
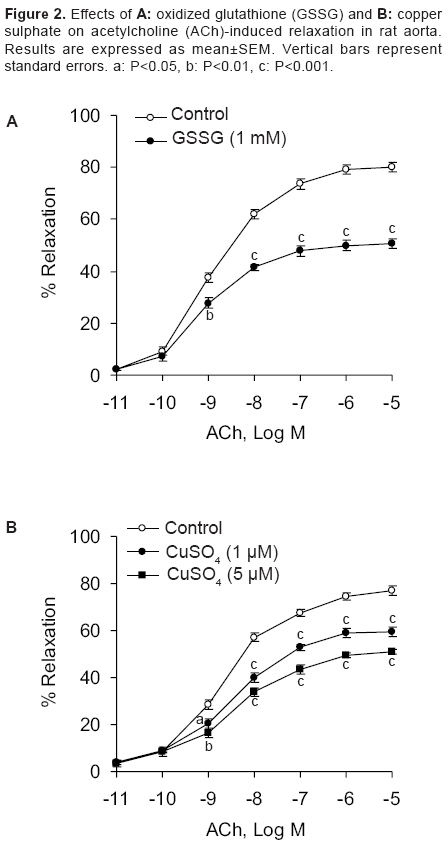
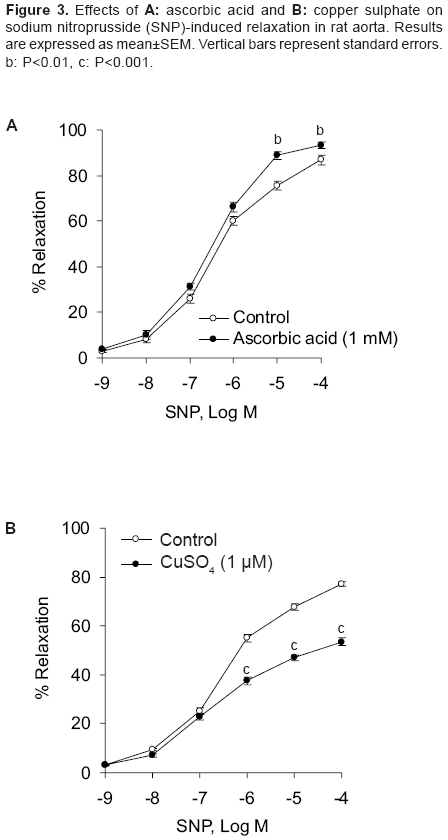
Indian Journal of Pharmacology, Vol. 38, No. 2, March-April, 2006, pp. 125-130 Research Paper Effect of redox agents on the response of rat aorta to nitric oxide and sodium nitroprusside Sardar KK, Sarkar SN, Bawankule DU, Mishra SK, Raviprakash V Division of Pharmacology and Toxicology, Indian Veterinary Research Institute, Izatnagar - 243 122, UP Code Number: ph06031 Abstract Objective: To study the redox regulation of vascular responses to endogenous nitric oxide (NO) and NO derived from nitrovasodilator sodium nitroprusside (SNP) in isolated rat aorta.Materials and Methods: To determine the influence of reducing [ascorbic acid (1 mM) and reduced glutathione (GSH) (1 mM)] and oxidizing agents [oxidized glutathione (GSSG) (1 mM) and CuSO4 (1 and 5 µM)] on the vasodilation caused by acetylcholine (ACh; 10-11 -10-5 M) and SNP (10-9 -10-4 M). Isometric tensions were measured in isolated aorta by a force transducer and recorded in a computer, using Chart V4.1.2 software. Results: ACh and SNP produced relaxation of rat aortic rings that was dependent on concentration. The rings were preconstricted with L-phenylephrine (1 µM). It was observed that oxidizing and reducing agents caused opposite effects on vasodilation induced by NO in rat aorta. Ascorbic acid and GSH potentiated the responses to NO, causing a leftward shift in the concentration-response curve of ACh with significant increase in the pD2 and the Emax . GSSG and CuSO4 inhibited relaxation caused by ACh and shifted the concentration-response curve to the right. In concentration-responses induced by SNP, ascorbic acid significantly increased the pD2 and Emax values from 5.85 ± 0.08 to 6.24 ± 0.05 and 80.83 ± 1.37% to 89.26 ± 1.49%, respectively. However, CuSO4 significantly decreased these values from 5.85 ± 0.02 to 4.56 ± 0.10 and 77.18 ± 0.82% to 53.52 ± 1.60%, respectively. Potentiation of NO response by reducing agents may be related to either increased availability of nitroxyl anion (NO- ) or reduction in superoxide anion radical (O2 ·- ). The opposite could be true for the oxidizing agents. Conclusion: The findings of this study suggest that reducing agents like ascorbic acid can improve the vascular responses to NO under oxidative stress. Keywords: Reducing agents, vasodilators, vitamin C Introduction Nitric oxide (NO) is a potent vasodilator. It is synthesised endogenously by the vascular endothelium that plays an important role in the regulation of vascular functions. Endothelial dysfunction is associated with various vascular disorders like atherosclerosis, systemic and pulmonary hypertension, arterial thrombotic disorders, angina pectoris, and stroke.[1] NO is also generated from a number of clinically important compounds called nitrovasodilators. These nitric oxide donor drugs are used in the treatment of disease conditions related to NO deficiency, such as, angina pectoris and pulmonary hypertension. Cellular redox state is believed to be an important factor in determining response in vascular smooth muscle that is related to NO. In addition, it determines cellular responses and diseases that are induced by stress. Inherent in these responses are reactive oxygen species (ROS) that inflict direct cellular damage. Vascular disease due to impaired NO bioactivity is primarily attributed to superoxide anion radical (O 2·- ), which is capable of rapidly inactivating endothelium-derived NO.[2] Therefore, the aim of the therapeutic interventions is to increase the NO bioavailability either by increasing NO production or decreasing O 2·- generation in the endothelium. Oxidative stress has been identified as an important factor in the development of tolerance to organic nitrates.[3] Release of NO from the nitrovasodilators involves redox regulation via endogenous reductants and oxidants. Sodium nitroprusside (SNP), a multivalent anion and NO donor, requires 1 electron reduction to initiate NO release. However, the nitrovasodilator 3-morpholino-sydnonimine (SIN-1) can spontaneously release NO by undergoing 1 electron oxidation.[4] In the living system, NO can exist in a variety of redox forms, such as, nitrosonium cation (NO+ ), NO free radical (NO· ), and nitroxyl anion (NO- ) depending on the source of the NO. It is not known which among these forms is responsible for vascular relaxation. Evidences suggest that NO- is more physiologically relevant.[5] However, according to Dierks and Burstyn,[6],[7] NO· is the only redox state that can activate soluble guanylyl cyclase (sGC). If it is so, the relaxant activity of NO- should be due to its oxidation to NO· . This was contradicted by the finding that NO- from Angeli′s salt (Sodium trioxodinitrate, Na 2 N 2 O 3 ) could mediate vascular relaxation without conversion of NO- to NO· , and this relaxation was mediated through GC activation.[8],[9] NO is believed to cause relaxation of vascular smooth muscle by activation of sGC and consequent rise in intracellular cyclic GMP. Activation of sGC can be accomplished with NO donors.[10] Redox active agents can alter the activity of sGC.[7],[11] Thiol reductants like dithiothreitol and reduced glutathione (GSH) cause inhibitory as well as stimulatory effects on sGC, while thiol oxidant, such as, oxidized glutathione (GSSG) inhibits the activity of sGC. Available literature reveals the importance of cellular redox state or processes in the regulation of the activity of sGC and the response in vascular smooth muscle that is related to NO. As both endogenous NO and NO donors primarily act through stimulation of sGC, it is important to examine the effects of redox reagents on the vasodilator responses elicited by NO. Several underlying signaling processes in vascular dysfunction are influenced by alterations in the status of intracellular redox. A better understanding of the regulation of function of vascular smooth muscle cell will provide further insight into the pathophysiological mechanisms that contribute to vascular changes and end-organ damage associated with hypertension. It could permit identification of potential novel therapeutic targets in the prevention and management of vascular disorders. Oxidative stress plays an important role in the dysfunction of endothelium and development of atherosclerosis. Modification of vascular risk factors and employment of antioxidants have been shown to improve endothelial function. In rat coronary artery, redox compounds have been shown to influence nitrovasodilator induced relaxation in vitro .[12] In patients with coronary artery disease, where vasodilator responses to acetylcholine (ACh) and SNP were reduced, ascorbic acid produced a beneficial effect.[13] Anderson et al .[14] also demonstrated the efficacy of antioxidants in the therapy of coronary artery disease. Ulker et al .[15], [16] showed that antioxidants, such as, ascorbic acid and tocopherol protected hypertension associated with enhanced oxidative stress. As information on the cellular redox state in influencing vasodilation of rat aorta by NO is scanty, we evaluated the effects of different reducing and oxidizing agents on the relaxation induced with ACh and SNP in rat aorta. Materials and Methods Chemicals Animals Recording of tension in aortic rings Assessment of redox regulation in rat aorta Statistical analysis Results
Effect of reducing agents on responses in rat aorta induced by ACh Effect of oxidizing agents on responses in rat aorta induced by ACh Effect of ascorbic acid and CuSO 4 on responses in rat aorta induced by SNP Discussion This study reports the redox regulation of vasodilator responses to NO in rat thoracic aorta. Influence of reductants, such as, ascorbic acid and GSH and oxidants like CuSO 4 and GSSG on relaxation produced by endogenous NO (released by ACh-induced endothelial stimulation) and by NO released from NO donor compound (SNP) was evaluated in rat aorta model. The important observations of this study are that oxidizing and reducing agents had opposite effects on vasodilation induced by NO in rat aorta. The relaxant responses to endothelium derived nitric oxide (EDNO) and SNP were potentiated by the reductants, ascorbic acid and GSH in rat aorta. Oxidants CuSO 4 and GSSG markedly inhibited vasodilator responses to ACh in rat aortic rings with intact endothelium and the responses to nitrovasodilator, SNP in rings denuded of endothelium. NO can exist in three different interrelated redox forms, i.e., NO+, NO· , and NO- with distinct properties and biological functions.[18],[19] The NO donor, SNP is known to contribute primarily NO+ species. The mechanism by which ascorbic acid potentiated the responses to EDNO and SNP is not clear, but may be related to either scavenging of intracellular O 2·- or greater availability of NO- redox species. Exogenous GSH had an effect similar to that of ascorbic acid, but it is widely believed that plasma membrane is impermeable to extracellular GSH.[20] Studies with vascular endothelial cells have, however, shown significant increases in the intracellular concentration of GSH in the presence of extracellular GSH.[21],[22] Findings of this study with ascorbic acid are consistent with the previous reports, wherein this compound was shown to improve endothelium-dependent flow-mediated dilation in patients with coronary artery disease[23] and lower pressure in hypertensive subjects.[24] Inactivation of EDNO by reactive oxygen species (ROS), viz., O 2·- may inactivate NO to produce vascular dysfunction. The observations of this study with ascorbic acid in rat aorta suggest that the antioxidant may improve rat aortic circulation in the event of an oxidative stress. Several reports reveal that ascorbic acid has variable effects on NO/nitrovasodilator-induced vasodilation in various blood vessels. Ascorbic acid reduced the responses to EDNO and authentic NO in rabbit aortic rings. In the same preparation, it potentiated relaxation to nitroprusside and S-nitroso-N-acetyl penicillamine (SNAP), but again inhibited relaxation to GTN.[25] In rat coronary artery, ascorbic acid potentiated the relaxant responses to authentic NO, but inhibited the vasodilator responses of 3-morpholino-sydnonimine (SIN-1) and SNAP.[12] Potentiation of the responses to EDNO and SNP by ascorbic acid and GSH in rat aorta possibly relates to a common mechanism of potentiation by these two reducing agents. As discussed elsewhere, among the three redox species of NO, nitroxyl anion (NO- ) is physiologically the most relevant one. This is because it is produced in abundance, endogenously.[8],[26] It is possible that NO+ derived from SNP undergoes reduction in the presence of reducing agents to form NO- . This could be the possible mechanism of potentiation of the nitrovasodilator responses by the reducing agents. In our laboratory, contribution of Na+ -K+ -ATPase to the potentiation of NO responses to reducing agents was evaluated by assaying the effect of ascorbic acid and GSH on 86 Rb-uptake in pulmonary artery of sheep. It was found that neither ascorbic acid nor GSH had any significant effect on either basal or SNP stimulated 86 Rb-uptake (unpublished data). This observation suggests that membrane bound Na+ pump may not contribute to the potentiation of NO responses induced by ascorbic acid. CuSO 4 and GSSG could reduce the availability of NO- species and therefore, inhibit relaxant responses of SNP in rat aorta. Current findings with CuSO 4 are at variance with the finding in rabbit aorta. De Saram et al .[25] reported that CuSO 4 had no significant effect on relaxation to nitroprusside in rabbit aortic rings. CuSO 4 has, however, been shown to reduce ACh-induced relaxation in rat aorta.[27],[28] Copper has been shown to increase the tissue cyclic GMP level in rat pulmonary artery and to stimulate NO synthase activity.[29] These mechanisms, however, do not explain the inhibitory effect observed in rat aorta. It is quite possible that oxidizing agents, viz., GSSG and CuSO 4 may reduce the availability of NO- species for the vasodilator responses of SNP in rat aorta. The other possibility of copper inhibiting nitrovasodilator responses may relate to O 2·- generation by this transition metal, which reacts with NO to produce peroxynitrite (OONO- ). Endothelial production of O 2·- by copper has been reported to cause altered NO activity and endothelial dysfunction.[28] Both GSH and GSSG have been shown to produce coronary vasodilation via NO and sGC-dependent mechanism in rats.[30] Although the present investigation has not made an attempt to define responses by GSH and GSSG that are dependent on endothelium, it is less likely that they alter the vasodilator responses to EDNO and SNP in rat aorta through endogenous NO formation/cGMP formation. This is evident from the findings that GSH and GSSG had opposite effects on the relaxation mediated by NO in rat aortic rings. Furthermore, these two compounds had no significant effect on the contraction elicited by submaximal concentration of phenylephrine in rat aortic rings. It is well documented that basal NO production has significant inhibitory influence on contraction elicited by vasoconstrictors. Therefore, substances modifying basal NO release would modify the responses to vasoconstrictors like phenylephrine. The contribution of endogenous NO or sGC pathway is not evident in rat aorta in the presence of GSH. Most likely, the oxidant and the reductant properties of GSSG and GSH, respectively, determine the modulation of NO responses in rat aorta. Based on the findings of this study, it may be concluded that oxidants cause inhibitory effect, while reductants produce an opposite effect on the vasorelaxation induced by ACh (EDNO) and NO donor - SNP in rat aorta. However, further studies are required to define their mechanisms of action. Acknowledgments The authors thank the Director, IVRI, Izatnagar, for providing financial assistance and necessary facilities to conduct this study.References
Copyright 2006 - Indian Journal of Pharmacology The following images related to this document are available:Photo images[ph06031f1.jpg] [ph06031f3.jpg] [ph06031f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}