 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Indian Journal of Pharmacology, Vol. 38, No. 4, July-August, 2006, pp. 266-270 Research Paper The effect of Hypericum perforatum extract against the neurochemical and behavioural changes induced by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) in mice M. Mohanasundari, S. Sethupathy# , M. Sabesan Department of Zoology, Annamalai University, Annamalainagar-608 002, #Division of Biochemistry, RMMCH, Annamalai Univeristy, Annamalainagar-608 002, Tamil Nadu, India Code Number: ph06070 Abstract Objective: Hypericum perforatum extract (HPE), known for its antidepressant effect, has been explored in the present study for its protective role against MPTP induced neurotoxicity.Materials and Methods: Mice were treated with 20 mg/kg of MPTP, four injections i.p., at 2 h intervals within 24 h. HPE was administered at different doses of 100, 200 and 300 mg/kg (p.o) in different groups once a day for seven days and the dose on the first day was given 30 min prior to first MPTP injection. Striatal dopamine (DA) and its metabolites, antioxidant status were analysed. The behavioural changes were studied using the rotarod test, hang test and narrow beam test. Results: HPE significantly ( P <0.05) improved the behavioural activities, striatal neurotransmitter levels and striatal antioxidant status in a dose dependent manner and significantly ( P <0.05) reduced TBARS levels. Conclusion: HPE possesses significant antioxidant activity and renders neuroprotection which was more pronounced at the dose of 300 mg/kg against MPTP induced neurotoxicity. Keywords: Neuroprotection, Parkinson's disease, St. John's wort. Introduction Hypericum perforatum, commonly called St. John's wort, has been used for centuries as a medicinal herbal remedy. The extract of the herb is widely prescribed for the treatment of mild to moderate depression, outselling fluoxetine by a factor of four and its popularity is growing world wide.[1] Parkinson's disease (PD) is a common neurodegenerative disorder characterised by a progressive loss of dopaminergic neurons in susbtantia nigra. Exposure of experimental animals to 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine (MPTP) provides a valuable model of neurotoxicity with behavioural, pathological and neurochemical features remarkably similar to those of PD. L-dopa is the most efficacious antiParkinsonian drug and virtually all patients respond to its administration. However, a major limitation to the chronic use of L-dopa is the develop-ment of motor complications such as motor fluctuations and dyskinesia.[2] With advancing disease, it becomes increasingly more difficult to deliver a dose of L-dopa that both controls Parkinsonian motor features and avoids dyskinesia.[3] Thus, neuroprotective strategies employing antioxidant is a good approach in improving the treatment of PD.[4],[5],[6] There is a need to search for alternative medicine, with neuroprotective effects, without side effects, for the management of PD. It has been recently reported that the effect of the H. perforatum extract might be mediated by an increase in brain levels of serotonin (5-HT), dopamine (DA), norepinephrine or by stimulation of opioid receptors in the central nervous system (CNS).[3],[7] The H. perforatum extract contains many phenolic compounds, namely flavonoids and phenolic acids, suggesting that they could have important antioxidant, antiinflammatory properties.[8] The polyphenols have the ability to penetrate the blood-brain barrier by simple diffusion which fulfill the requirements for a potential neuroprotective agent.[9] So, the present study has been designed to evaluate the dose dependent effects of HPE against MPTP induced neurotoxicity in mice. Materials and Methods Animals Male Swiss albino mice ( Mus musculus ), weighing 25-35 g, were procured from the central animal house, Department of Experimental Medicine, Rajah Muthiah Medical College, Annamalai University, Annamalainagar. The animals were maintained at the central animal house and were fed on a standard balanced diet (Hindustan Lever, Bangalore) and provided with water ad libitum . All studies were conducted in accordance with the National Institute of Health Guide. Chemicals MPTP (salt) was obtained from Sigma Chemical Co. USA and all other chemicals used were of analytical grade. Plant material and preparation of extracts H. perforatum plants were collected from Western Ghats of Nilgiris, Tamil Nadu and were botanically authenticated by the Department of Botany (Voucher Specimen Ac No. 2456), Annamalai University, Annamalainagar, Tamil Nadu. The leaves were air dried at room temperature, finely powdered with auto-mix blender and stored in a deep freezer until the time of use. The methanolic extract prepared using Soxhlet was concentrated by rotary evaporator at 40°C and stored in a cool place. Experimental design The animals were divided into six groups, each consisting of six mice. Group I served as vehicle control (10% Tween 80, p.o.), Group II received 300 mg/kg HPE (p.o.), Group III received 20 mg/kg MPTP four injections at 2 h intervals, i.p .,[10] Groups IV, V and VI received HPE at the dose of 100, 200 and 300 mg/kg (p.o.), respectively. The treatment was given on the initial day, 30 min. prior to first injection of MPTP and once a day for another six days of the experimental period. At the end of experimental period (after 7 days of treatment) the animals were fasted overnight and sacrificed by cervical decapitation before stunning. The brains were excised immediately, the striatal region was identified using the stereotaxic atlas of an albino mouse brain.[11] The fresh brain was serially sectioned and the striatum was separated. The tissue was homogenised in ice cold 0.1% M PBS solution and used for further analysis as mentioned later. Motor integration tests were assessed by different methods Rotarod test Motor coordination was measured on the seventh day using an automated rotarod (Amni, Rotar Instrumentation, Columbus, OH, USA). The animals were exposed to 10 trials on a rotating rod at 10 rpm at 5 min. intervals with a cut off time of 180 seconds.[12] The rotor was divided into two compartments, which could allow two mice at a time. The average retention time on the rod was calculated. Hang test Neuromuscular strength was determined on the seventh day in the grid hang test. The mice were lifted by their tails, slowly placed on a horizontal grid (grid 12 cm 2 opening 0.5 cm 2) and supported until they grabbed the grid with their fore and hind paws. The grid was then inverted so that the mice were allowed to hang upside down. The grid was mounted 20 cm above a hard surface to discourage falling, but not leading to injury in case of a fall. The apparatus was equipped with a 3-inch wall to prevent the animals from transversing to the upper side of the grid. The animals were required to stay on the grid for 30 seconds and 10 chances were given with 1 min intervals and the maximum hanging time was recorded.[13] The calculation was made as percentage of hanging time
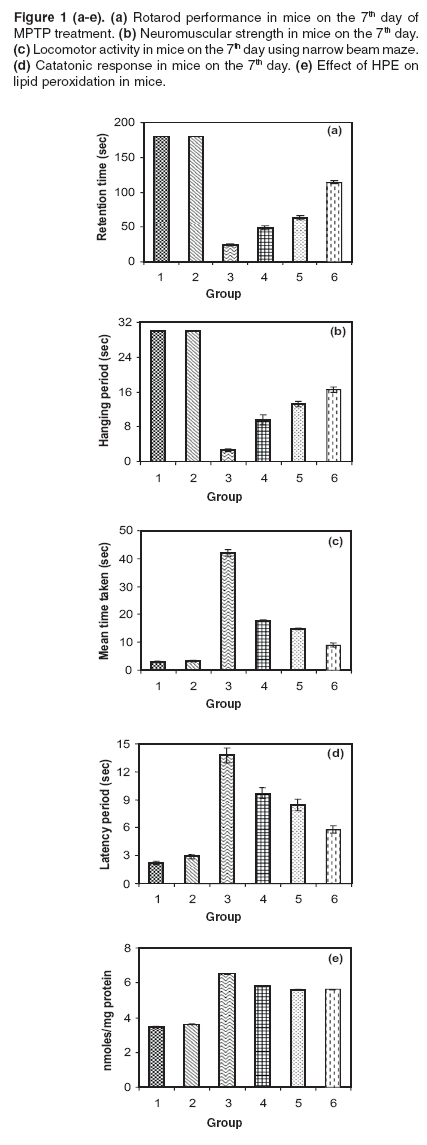
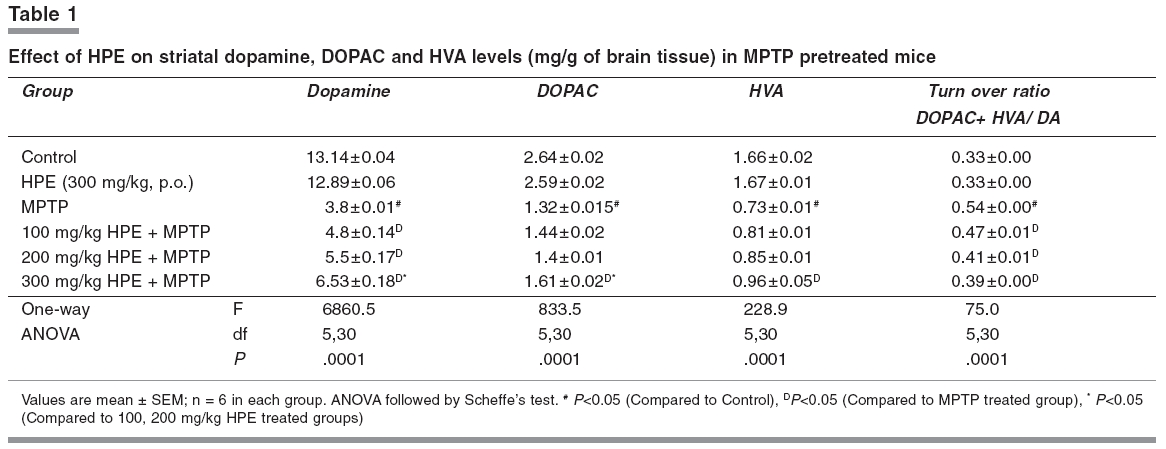
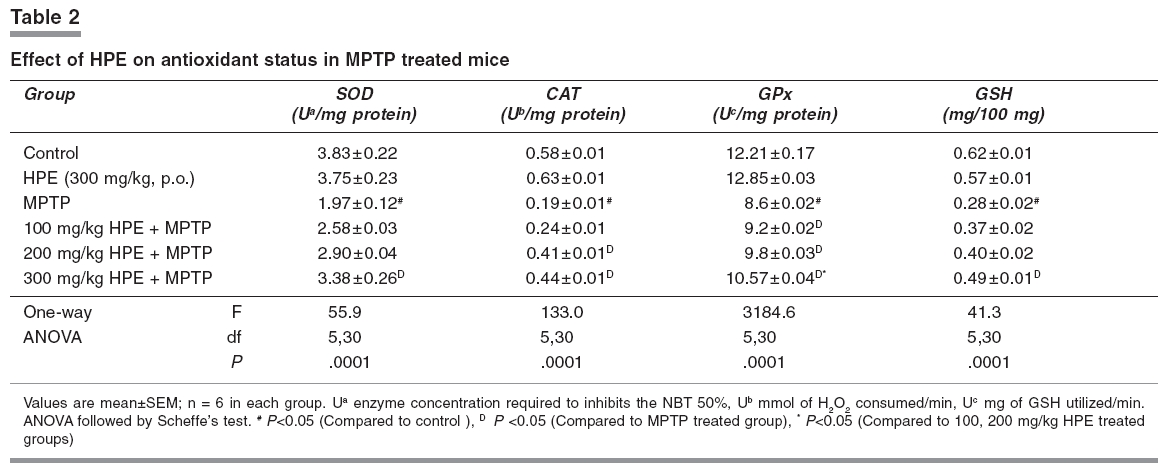
Narrow beam maze (NBM) This maze consisted of a start-and-goal-box and the runway was made of a smooth wooden narrow beam of 180 cm length with a runway width of 2 cm. The beam was elevated off the ground by 1 meter with additional supports. The inner diameter of start-and-goal-box was 13 cm width and 40 cm high walls and the inner walls were painted black. The animals were trained in an NBM for 10 trials per day with 1 min interval. The semi-starved mice were allowed to explore the NBM for 10 trials with 1 min interval on the seventh day of experimental period and during this exploration animals were motivated and rewarded with food pellets in the goal box. The journey time between start-box-and-goal-box was measured.[14] Catatonic behaviour Catatonia was measured by placing the animals on a flat horizontal surface with both hind limbs on a square wooden block (3 cm height): latency to move was calculated in seconds.[15], [16] Biochemical analysis Measurement of dopamine and DOPAC The brain samples were weighed immediately after dissection, frozen and stored at -80°C until assay. The striatal samples were sonicated in ice-cold 0.2 M perchloric acid containing 100 mg/ml, isoproterenol as internal standard. Homogenates were centrifuged at 3000 rpm for 15 min at 4°C. The supernatant was filtered and an aliquot (30 ml) was used for the determination of dopamine and DOPAC levels using HPLC with an electrochemical detector ([ECD]; Eicom, Kyoto, Japan). The amounts of dopamine and DOPAC were calculated by comparing the simple peak with the internal standard peak and expressed as microgram per gram of tissue weight.[17] Estimation of lipid peroxidation products Thiobarbituric acid was added to the brain homogenate under acidic conditions and the absorbance of colour that developed after heating was estimated spectrophotometrically at 535 nm.[18] Assay of superoxide dismutase (SOD EC.1.15.1.1) SOD was assessed by the inhibition of formation of NADH-phenazine methosulphate nitroblue tetrazolium formazon.[19] The reaction was initiated by the addition of NADH after incubation for 90s and stopped by the addition of glacial acetic acid. The colour formed at the end of the reaction was extracted into the butanol layer and measured at 520 nm. Assay of catalase (CAT, EC.1.11.1.6) CAT was assayed colorimetrically as per the method of Sinha.[20] Dichromate in acetic acid was converted to perchromic acid and then to chromic acetate when heated in the presence of H 2 O 2 . The chromic acetate formed was measured at 620 nm. The catalase preparation was allowed to split H 2 O 2 for different periods of time. The reaction was stopped at different time intervals by the addition of a dichromate-acetic acid mixture and the remaining H 2 O 2 was determined colorimetrically as chromic acetate. Assay of glutathione peroxidase (GPx EC.1.11.1.9) GPx was estimated as described by Rotruck et al.[21] A known amount of brain homogenate was allowed to react with H -2 O 2 in the presence of GSH for a specified time period, then the remaining GSH was allowed to react with DTNB and the developed yellow colour was measured at 412 nm. Estimation of reduced glutathione GSH in brain homogenate was measured according to the method of Ellman.[22] This method is based on the development of a yellow colour when 5,5'-dithio-bis-2-nitrobenzoic acid (DTNB) is added to compounds containing sulfhydryl groups. Statistical analysis One way analysis of variance followed by Scheffe's test was employed for the analysis of behavioural variances and neurochemical parameters. P < 0.05 was considered significant. Results [Figure - 1]a depicts the results of the rotarod test. The retention time significantly decreased in the MPTP treated group when compared with the control group and it was significantly improved ( P < 0.05) in 100, 200 and 300 mg/kg of HPE groups. Among HPE groups, 300 mg/kg showed marked improvement ( P < 0.05) compared to 100 and 200 mg/kg. [Figure - 1]b depicts the results of the hang test. The hang time significantly decreased ( P < 0.05) in the MPTP treated group compared to the control group. The hang time significantly ( P < 0.05) improved in 100, 200 and 300 mg/kg HPE treatment groups. Again in HPE groups, 300 mg/kg showed marked improvement ( P < 0.05) in hanging time compared to 100 and 200 mg/kg. [Figure - 1]c shows the results of the narrow beam test. The crossing time of 180 cm length wooden path significantly ( P < 0.05) decreased in the MPTP treated animals as compared to the control group. The HPE groups 100, 200 and 300 mg/kg showed marked improvement in locomotor ability as compared to the MPTP treated group. The 300 mg/kg HPE group showed a significant improvement in the locomotor ability compared to lower doses. [Figure - 1]d reveals the catatonic response. The latency period was significantly ( P < 0.05) increased in the MPTP treated animals as compared to the control group. The 300 mg/kg HPE group showed a significant ( P < 0.05) reduction in the latency period compared lower doses. [Table - 1] displays the striatal dopamine and the metabolite levels in the control and experimental groups. Dopamine and its metabolite DOPAC and HVA markedly ( P < 0.05) decreased in the MPTP treated animals while their levels significantly ( P < 0.05) improved in 300 mg/kg HPE treated animals when compared to 100 and 200 mg/kg HPE treatment groups. Dopamine turnover ratio significantly ( P < 0.05) increased in the MPTP group and significantly decreased ( P < 0.05) in all HPE treatment groups, wherein with 300 mg/kg HPE, the decrease was marked. Striatal TBARS and antioxidant status are presented in [Table - 2] and [Figure - 1]e. Striatal TBARS was significantly ( P < 0.05) elevated in the MPTP group compared to the control group. HPE, 300 mg/kg, significantly ( P < 0.05) decreased the TBARS level compared lower doses. SOD, CAT, GPx and GSH activities were significantly decreased ( P < 0.05) in MPTP treated animals as compared to the control group. These activities were restored significantly ( P < 0.05) in 300 mg/kg HPE group. Discussion MPTP caused significant overt and subtle behavioural manifestations. The study also reveals the existence of quantitative behavioural responses within a short span of seven days in MPTP-lesioned animals to the administration of HPE in a dose dependent manner. MPTP administered mice, subjected to the rotarod test, revealed a significant loss of muscular coordination and the poor performance in the hang test. This could be due to loss of muscular strength as reported earlier.[13] The narrow beam maze, used to test the balance and working performance,[14] was significantly altered by MPTP treatment. MPTP also induced the catatonic response. The striatum is known to be involved in head and forelimb motor control. Altered behavioural responses in mice, following DA depletion or striatal damage, is considered to be similar to human Parkinsonism.[23],[24],[25],[26] In the present study, a significant reduction of striatal DA and its metabolites in MPTP treated mice was observed. The dopamine turnover increased after MPTP intoxication in mice. This increased turnover ratio is generally considered as a compensation for dopamine depletion. Increased striatal lipid peroxidation with decreased antioxidant status was observed in MPTP treated animals. HPE prevented the motor impairment in a dose dependent manner and also decreased the latency period of catatonic behaviour, altered by MPTP. The animals which were administered 300 mg/kg HPE had a significant improvement than those which received lower doses. The loss of striatal dopamine was at least partially prevented when mice were administered HPE at various doses and the beneficial effect was maximum at 300 mg/kg than with other doses. Recently, hyperforin, the fluoroglucinol derivative of HPE has gained interest, as it represents the major component responsible for the inhibition of the uptake of several neurotransmitters in vitro.[27] HPE as well as hyperforin treatment modulates neurotransmitter levels in the brain of rodents.[28] In addition, there are several reports on HPE inhibiting dopamine reuptake.[29],[30] Due to this mechanism, the dopamine level was retained in the synaptic region. It has been reported that MAO-B inhibition was evident with H.perforatum extract fractions containing higher concentrations of flavonoids.[31],[32] Possibly due to inhibition of MAO-B, conversion of MPTP to MPP+ is decreased resulting in decreased lipid peroxidation. In the present study, HPE reduced the MPTP induced lipid peroxidation (TBARS) in a dose dependent manner. Silva et al.[8] also demonstrated a significant protection against ascorbate/ Fe 2+ induced lipid peroxidation by ethanolic extract of HPE, in rat cortical synaptosomes. Hunt et al.[30] evaluated the antioxidant effect of HPE against the superoxide anion and demonstrated that a free radical-scavenging effect for most of the concentrations tested. They postulated that this effect could be related to its content in hypericin. In the present study, HPE effectively improved the antioxidant enzyme activities such as SOD, CAT and GPx in MPTP treated animals especially at the dose of 300 mg/kg. So, it is concluded that HPE can exert a significant neuroprotective effect and its usefulness in the management of Parkinson's disease needs exploration. References
Copyright 2006 - Indian Journal of Pharmacology The following images related to this document are available:Photo images[ph06070t1.jpg] [ph06070f1.jpg] [ph06070t2.jpg] |
| |||||||||

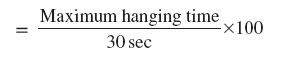
{kind=link}
{kind=link}
{kind=link}