 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
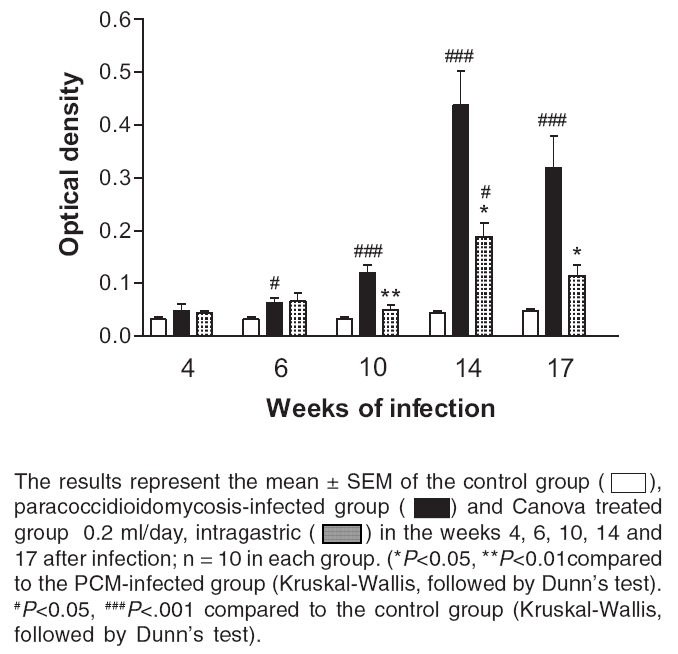
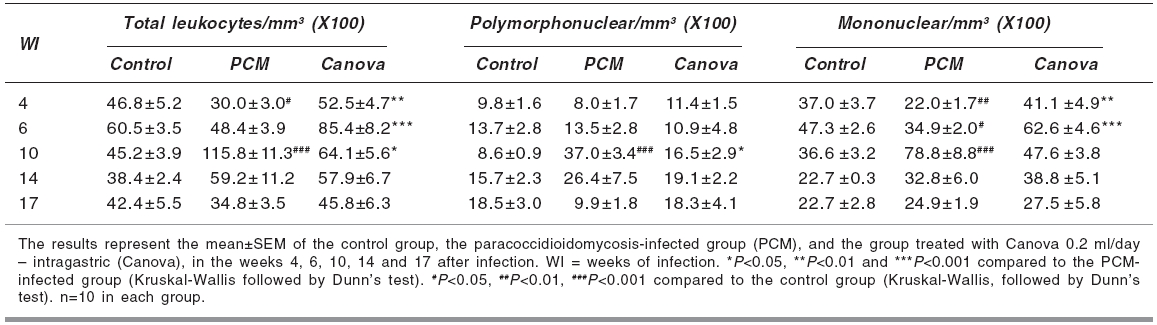
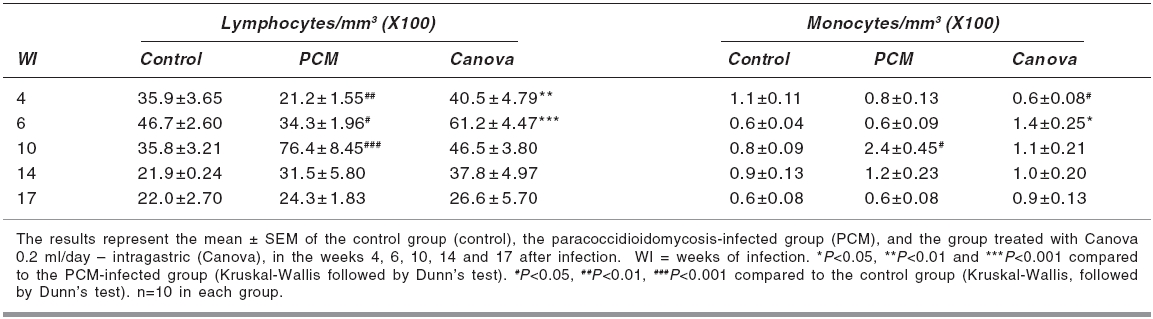
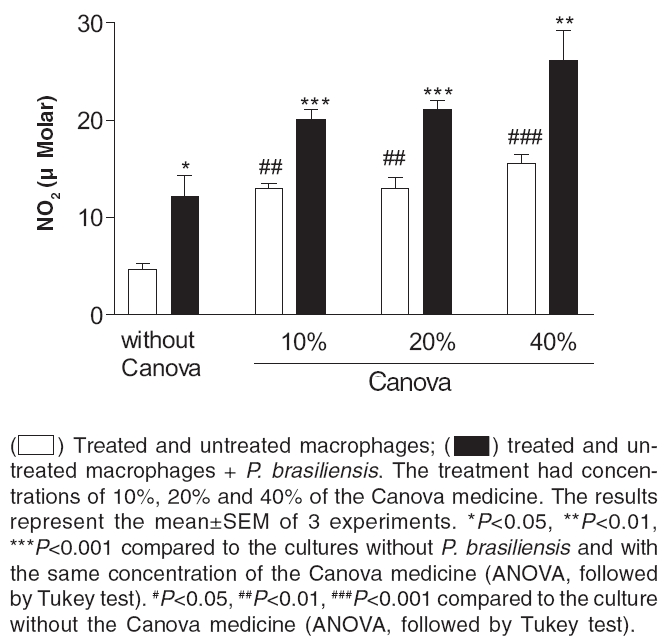
Indian Journal of Pharmacology, Vol. 38, No. 5, September-October, 2006, pp. 350-354 Research Paper In vivo and in vitro effects of the Canova medicine on experimental infection with Paracoccidioides brasiliensis in mice Takahachi G, Maluf MLF, Svidzinski TIE, Dalalio MMO, Bersani-Amado CA, Cuman RKN Department of Pharmacy and Pharmacology, State University of Maringa Date of Submission: 20-Jul-2006 Code Number: ph06097 Abstract Objective: To evaluate the in vivo and in vitro activity of Canova in experimental infection with Paracoccidioides brasiliensis.
Keywords: Immunomodulator, macrophage activation, systemic mycosis. Introduction The Canova ® (Canova of Brazil) method medicinal product is composed of 5 traditional homeopathic substances: Aconitum, Arsenicum album , Bryonia alba , Lachesis muta and Thuya occidentalis . This product has been used for patients with AIDS and cancers, where the immunological system is depressed.[1] Clinical and experimental observations demonstrate that the Canova medicine promotes an improvement in concomitant infectious and inflammatory conditions, apart from the reduction and regression of tumour[1],[2] in some cases. Experimental data demonstrates that the mechanism of action of the Canova compound is probably due to the increase in the immune response through macrophage activation.[3] Other actions such as a decrease in the production of the tumour necrosis factor-a (TNF-a),[3] stimulation of leishmanicidal activity and nitric oxide (NO) production in macrophages[4] are also attributed to Canova. Paracoccidioidomycosis (PCM) is a progressive, systemic fungal disease of granulomatous origin. It affects mainly the lungs, ganglions and mucosa, as well as other organs and systems with varying clinical signs and symptoms.[5] The disease is geographically limited to Latin American countries and some regions of Brazil are considered as reservoirs and endemic.[6],[7] In PCM, the effective and protective response of the host depends primarily on cell-mediated immunity, with neutrophils and macrophages playing an important role in phagocytosis.[8] High levels of anti P. brasiliensis antibodies are associated with the most severe forms of the disease.[9] The immunological dysfunction, associated with PCM, is characterised as a specific antigen defect of the cellular immunity which is reversible with the treatment of the disease and is possibly related to a Th2 standard immune response during the active disease.[10] The analysis of the cytokines production and immunopathological characteristics, in resistant and susceptible mice, indicate that resistance to P. brasiliensis infection is associated with an early activation of immune mechanisms which leads to a Th1 response. Susceptibility is related to the early production of mediators which inhibit the production of interferon gamma (IFN-g), which suppress the progress of the protective Th1 immunity.[11] The importance of the immunoregulatory effects of the cytokines in the resistance mechanism of PCM was demonstrated by in vitro and in vivo studies. It was shown that IFN-g produced during the cellular immune response activates macrophages which are important in controlling P. brasiliensis infection, thus proving the efficacy of the immunomodulatory therapy.[12],[13] In the present study, the effect of the Canova medicine on the evolution of experimental P. brasiliensis infection and on the activation of infected macrophage in vitro was evaluated. Materials and Methods Fungus The isolate P. brasiliensis Pb18 from the Pathology Department, Medical Faculty, State University of Sao Paulo (UNESP - Botucatu Campus) was used. The yeast cells were cultured in YPD (Yeast extract, Casein-peptone, Dextrose) semi-solid medium at 35°C for 7 days. To prepare the inoculum, the cells were washed 3 times with sterile phosphate buffer solution (PBS) of pH 7.2. The cell viability was determined, using the Janus Green vital stain. Cultures with a minimum of 80% viability were used and the concentration adjusted by counting in a Neubauer chamber. Animals Male Swiss albino mice, 4-6 weeks' old, from the Central Animal Facilities of the State University of Maringa were used. The animals were divided into 3 groups of 50 mice each: Canova (infected and Canova treated); PCM (infected but not treated) and Control (not infected and not treated). The mice were infected with 0.5 ml of the fungus suspension at a concentration of 1.0 X 10 7 cells/ml (5 X 10 6 yeast cells) i.p.[14] During the experimental period, the animals were housed under standard laboratory conditions with ad libitum water and balanced food. The experiments were approved by the University Ethics Committee of the State University of Maringα. Treatment The Canova ® medicine was administered intragastrically (0.2 ml/day).[4] The treatment was started on the same day of induction of infection and maintained for 17 weeks. Treatment evaluation During weeks 4, 6, 10, 14 and 17 after the infection was induced, 10 mice from each group were anaesthetised (50 mg/kg ketamine hydrochloride and 50 mg/kg thiazine hydrochloride, i.p.) and blood samples collected to determine the number of total and differential leukocytes and the total antibody titre against P. brasiliensis . A Neubauer chamber was used for the global leukocyte count, while the differential count was done by optical microscopy, using blood smears stained with the Leishman stain. The serum levels of specific antibodies were determined by the ELISA method, using P. brasiliensis culture filtrate as the antigen.[7] Commercial antimouse total immunoglobulin peroxidase conjugate (Sigma ®) was used to detect the antigen-antibody reaction. Macrophage culture Peritoneal cells were extracted from healthy mice, by washing the peritoneal cavity with 5 ml of cold PBS. After that, the cells were re-suspended in RPMI 1640 medium supplemented with 10% fetal bovine serum, 2 mM L-glutamine, 40 µg/ml gentamicin, 20 mM HEPES, 2.5x10 -5sub M 2-mercapto-ethanol. For the macrophage culture, 1 ml of a 2 X 10 6 cells/ml suspension was cultured in 24 well plates (CORNING ®) for 2 h at 37°C and 5% CO 2 . After incubation, the culture supernatants were aspirated and the non-adherent cells removed. The monolayer of macrophages was incubated again, at 37°C and 5% CO 2 , for 21 h 3sub with complete RPMI containing 10%, 20% or 40% of the Canova medicine. Culture without Canova was used as control. Co-culture of macrophage and P. brasiliensis After the incubation period with Canova, the supernatants of the cultures were discarded and the macrophages challenged against 4 X 10 4sub (1:50 fungus:macrophages) yeast cells of P. brasiliensis suspended in RPMI supplemented with 10% fresh mice serum. The co-cultures were re-incubated at 37°C and 5% CO 2 for 4 h. After incubation, the culture supernatants were removed and the nitric oxide level estimated. Cultures containing P. brasiliensis yeast cells or non-treated macrophages plus yeast cells were used as controls. Nitric oxide (NO) determination The NO 2 concentration, in the culture supernatants, was used as an indicator of NO generation, and was measured using the Griess reagent (1% sulphanilamide, 0.1% naphthylenediamine, 2.5% H 3 PO 4 ).[15] Plates of 96 wells (CORNING ®) were used and 50 µl of the supernatant was added to the same volume of the Griess reagent and incubated at room temperature for 10 min. After incubation, the absorbance was determined in a microplate reader (FL 600 BIO-TEK ®) using 530 nm filter, against a blank (culture medium and the Griess reagent), under the same conditions as described before. The results were expressed as µM of NO 2 , using a sodium nitrite standard curve. All determinations were made in triplicate. Statistical analysis The program GraphPad Prism ® (version 3.0, GraphPad Software, Inc.) was used for the statistical analysis. The results are expressed as mean±SEM. The groups difference were analysed in each week using non-parametric analysis of variance (one-way ANOVA), followed by the Tukey test or Kruskal-Wallis test, followed by the Dunns test, for multiple comparisons. The level of significance used was 0.05. Results Antibody level P. brasiliensis infection induced a significant increase in antibody level after the 6th week of fungal inoculation. A significant reduction of anti P. brasiliensis antibody was seen in the Canova-treated group compared to the PCM group in the 10th ( P < 0.01), 14th and 17th ( P < 0.05) weeks. [Figure - 1] Global and differential leukocyte count The infected, untreated mice initially presented a reduction in total leukocyte count in the 4th week of infection ( P < 0.05), due to the reduction in mononuclear cells ( P < 0.01), and lymphocyte count ( P < 0.05). In the succeeding weeks, the leukocyte count increased gradually with a peak during the 10th week of infection ( P < 0.001), with an increase in polymorphonuclear ( P < 0.001) and mononuclear cells ( P < 0.001). For the Canova-treated infected animals, this increase started in the 4th week ( P < 0.01), with a peak during the 6th week after infection ( P < 0.001). This increase was associated with an elevation in mononuclear cells ( P < 0.001), such as lymphocytes and monocytes. [Table - 1][Table - 2] Nitric oxide The Canova medicine caused an increase in nitric oxide in the macrophage culture supernatant, with all the concentrations tested, without any significant difference between them. The culture with Canova promoted a higher increase ( P < 0.001) in nitric oxide, than P. brasiliensis alone ( P < 0.05). The supernatant of the cultures containing P. brasiliensis and Canova in the tested concentrations were also determined and the concentrations were close to 1.0 µM. [Figure - 2] Discussion In PCM, the response of the host to the infective agent occurs primarily through cell mediated immunity. Neutrophils and macrophages have an important role in the phagocytosis of PCM.[8] However, in this systemic mycosis, depressed cell mediated immunity is commonly observed.[16] Antifungal therapy, combined with an immunomodulator, might be efficient in the treatment of this disease.[12] On the other hand, many works have been carried out to study the control of the PCM using antifungal drugs, so this effect is well estabilished. In this way, we did not use, in this protocol, an antifungal-treated group. Thus, the effect of the Canova medicine in experimental murine PCM was studied. The results of the present study revealed a significant decrease in total leukocytes, with a significant decrease in the lymphocyte count in the 4th week of PCM infection. In the 10th week of infection, a significant increase in the total leukocyte count was observed in the PCM-infected group, while in the Canova-treated animals, this elevation was earlier, starting in the 4th week and peaking in the 6th week, with an increase in the number of mononuclear cells. The global and differential leukocyte counts of the Canova-treated animals were similar to those of the uninfected animals (control group) in the majority of the weeks (14th and 17th) studied. These facts indicate that the Canova medicine may already be acting in the first week of treatment, possibly favourably affecting the immunological response. This is reflected in the significant reduction of antibody level, while the positive control group exhibited high levels of the same from the 10th week after infection, persisting in the 17th week. These results are in accord with an increase in the number of mononuclear cells, with a peak at the 10th week of infection. This increase might reflect an elevation of B lymphocytes, since there is an increase in the polyclonal B lymphocytes in more severe cases of PCM.[9] As high levels of antibodies are associated with more severe forms of the disease and used as indicator of clinical recurrence,[9] the present results suggest that PCM infection was less severe in the Canova-treated group. A significant increase in NO production was observed in the macrophage culture incubated with P. brasiliensis, compared to those without the fungus. These findings indicate probable macrophages and involvement of the nitric oxide pathway. Gonzalez et al ,[17] also observed an increase in the fungicidal capacity of peritoneal macrophages in mice activated with IFN-g on conidia and yeast cells of P. brasiliensis , and that the incubation of these cells with nitric oxide synthase inhibitors reverted this effect, suggesting that the increased fungicidal activity could be mediated by nitric oxide production. In our experiments, supernatants of macrophage cultures, infected or not, and incubated with Canova presented higher NO concentrations than the control without Canova. Our data indicate that Canova has a potential for macrophage activation as also suggested by Piemonte &Buchi.[3] One of the characteristics of activated macrophages is the increase in NO production.[18] NO possess antimicrobial activity; through reaction with super oxides, peroxy nitrite is produced, which is cytotoxic and important for the cellular immunity.[19],[20] Many in vitro and in vivo studies have demonstrated a direct correlation between the NO production and the antimicrobial activity of the macrophages in the presence of various microorganisms, such as Leishmania amazonensis,[4] Histoplasma capsulatum,[21] Trypanosoma cruzi [22] and Schistosoma mansoni .[23] The present results suggest that the control of the infection may be due to the activation of the cellular response in the animals treated with Canova. When the macrophages are activated, they may have additional effects on the fungus, through the release of pro-inflammatory cytokines. Further studies are needed to know more about the antimicrobial activity of Canova. Acknowledgments This work was partially supported by Conselho Nacional de Desenvolvimento Cientνfico e Tecnológico. We thank Dr. Roberto Piraνno for providing the Canova medication samples. We are also grateful to Juarez Antonio da Silva, Sandra Regina Corbelo Pereira and Eliana Guilhermetti for their technical assistance. References
Copyright 2006 - Indian Journal of Pharmacology The following images related to this document are available:Photo images[ph06097f1.jpg] [ph06097t1.jpg] [ph06097t2.jpg] [ph06097f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}