 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
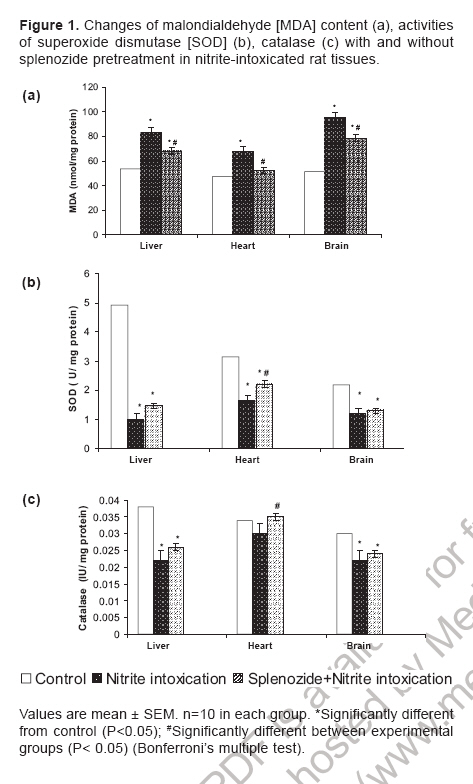
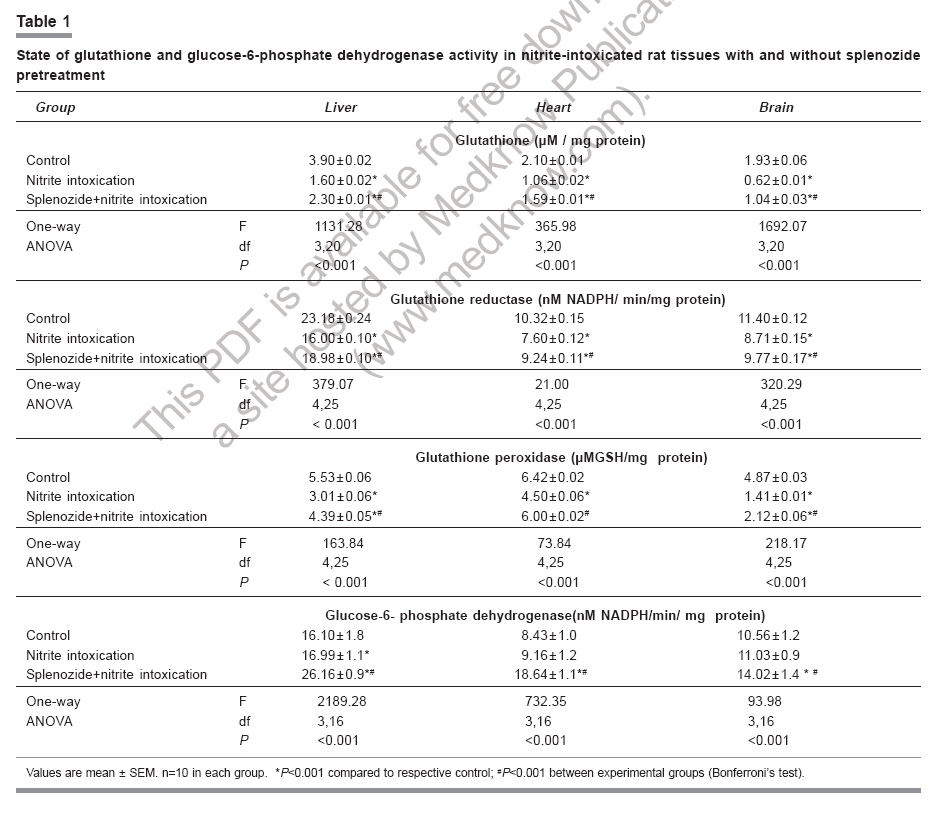
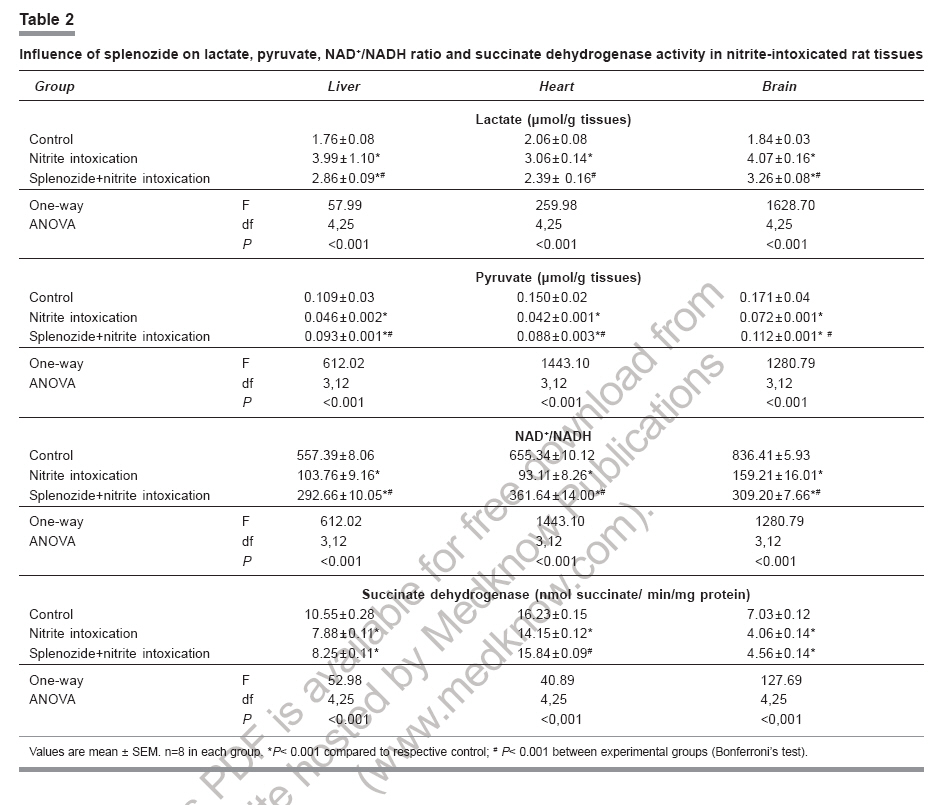
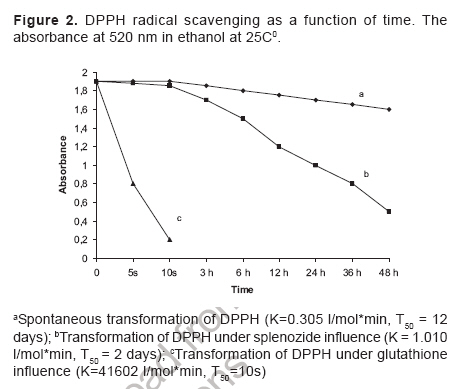
Indian Journal of Pharmacology, Vol. 38, No. 6, November-December, 2006, pp. 414-418 Research Paper Role of complex nucleosides in the reversal of oxidative stress and metabolic disorders induced by acute nitrite poisoning Gonchar O, Mankovskaya I, Klyuchko E Department of Hypoxic States, Bogomoletz Institute of Physiology National Academy of Sciences of Ukraine, Kiev Date of Submission: 31-Jan-2006 Code Number: ph06117 Abstract Objective: To investigate the possible mechanisms of splenozide influence on oxidative stress and metabolic disorders in tissues caused by acute nitrite intoxication in rats.Materials and Methods : Nitrite poisoning (NaNO2, 60 mg/kg, s.c.) was induced in rats pretreated with splenozide (natural metabolite, nucleoside complex; 3 mg/kg, i.p.). The parameters of lipid peroxidation (LPO), antioxidant defense system (superoxide dismutase [SOD], catalase [CAT], glutathione content [GSH], glutathione reductase [GR], glutathione peroxidase [GPx], glucose-6-phosphate dehydrogenase [G6PDH]), lactate and pyruvate concentrations, NAD+/NADH ratio in cytosol and mitochondrial succinate dehydrogenase (SDH) activity were evaluated in the liver, myocardium and brain. Results: Splenozide pretreatment decreased LPO, stimulated the G6PDH, GR and GPx activity, and increased the intracellular glutathione level in all studied tissues. Its effect on SOD, CAT and SDH activity depended on the type of tissue studied. Splenozide caused a reduction of lactate concentration and accumulation of oxidized NAD in cytosol. Conclusion: Splenozide demonstrated a weak scavenging activity against 1,1-diphenyl-2-picrylhydrazyl (DPPH) free radicals in vitro . Its antioxidant effect during nitrite intoxication may be due to the maintenance of glutathione recycling activity through the activation of NADPH-dependent reactions, redox state restoration and antiacidotic effect. Keywords: Antioxidant, splenozide, splenic factor. Introduction Nitrites are widely used in the manufacture of dyes, treatment of textiles, curing of meat and in contemporary agriculture,[1] as a result of which, nitrite intoxication is a major contibutor towards environmental pollution. Supernormal concentrations of nitrites in living organisms cause life-threatening states. This xenobiotic may interact with N-alkyl-amides to form N-nitroso carcinogens, which cause methemoglobinemia, disturbance of oxygen transport with attendant tissue hypoxia and other diseases.[1],[2],[3] The excess generation of reactive oxygen species (ROS) in an organism is one of the general non-specific mechanisms of damage caused by many environmental chemical pollutants. Nitrite (NaNO 2 ) in toxic doses has been reported to enhance lipid peroxidation (LPO) and cause changes in antioxidant enzyme activity.[3],[4] Agents like NADH, ascorbic acid, α-tocopherol and selenium have been reported to protect against acute and chronic nitrite poisoning.[4],[5],[6] Against this background, the present study investigated the activity of splenozide on the oxidative stress status and the metabolic disorders in the heart, liver and brain of rat caused by acute nitrite intoxication, as well as in vitro antiradical activity. Materials and Methods Chemicals Splenozide was obtained from Komissarenko Institute of Endocrinology and Metabolism. Thiobarbituric acid, 1,1-diphenil-2-picrylhydrazyl (DPPH), epinephrine, NADP, NADPH, glutathione, 5', 5'-dithiobis-(2-nitrobenzoic acid), glucose-6-phosphate, and lactate dehydrogenase were purchased from Sigma Chemical Co., USA. All chemicals used were of analytical grade. Animals and study design Wistar rats (220-260 g) were housed in plexiglas cages (four animals per cage) and maintained in an air-filtered and temperature-controlled (20-22°C) room, under artificial light/dark cycle of 12 h. They received a standard pellet diet and water ad libitum . Acute nitrite intoxication was induced by injecting aqueous NaNO 2 (60 mg/kg, s.c.). The rats were decapitated 50 min after the injection, when methemoglobin formation was at its peak, i.e. ~ 34%. Splenozide was dissolved in saline and injected i.p. (3 mg/kg) three times according to the following scheme: 2 days, 1 day, and 30 min before the nitrite poisoning.[10] The rats were divided into three groups of 18 animals each: Group 1 : Control Preparation of subcellular fractions Animals were euthanized by the method approved by the European Convention for Animal Defense (Strassbourg, France, 1985). After decapitation the liver, heart, and brain were immediately removed, washed in cold saline and homogenized in the ice-cold buffer (0.25 M sucrose, 10 mM Tris-HCl, 1 mM EDTA, pH 7.4) with a motor-driven glass Potter-type homogenizer. The homogenates were centrifuged at 1000 g for 10 min to remove the cell debris. Mitochondria were isolated at 9000 X G for 15 min and the pellets were washed twice under the same conditions. Mitochondria were resuspended in 0.25 M sucrose. The 9000 X G supernatant was centrifuged at 15000 X G for 30 min and the supernatant was used for assay of enzyme activity and LPO. All procedures were carried out at 0°-4°C. Mitochondria were stored at -20°C. Biochemical assays Lipid peroxidation was determined by measuring the malondialdehyde (MDA) concentration.[11] Superoxide dismutase (Cu, Zn-SOD; EC 1.15.1.1) and catalase (CAT; EC 1.11.1.6) activities were estimated by the methods of Misra and Fridovich[12] and Aebi.[13] Reduced glutathione (GSH) content was determined by the method of Sedlak and Lindsay.[14] Activities of glutathione reductase (GR; EC 1.6.4.2), glutathione peroxidase (GPx; EC 1.11.1.9), and glucose-6-phosphate dehydrogenase (G6PDH; EC 1.1.1.49) were measured as per the methods of Carlberg and Mannervik,[15] Olinescu and Nita[16] and Deutsch,[17] respectively. Succinate dehydrogenase (SDH; EC 1.3.99.1) activity was estimated according to the method of Veeger et al .[18] Contents of lactate and pyruvate were assayed on the liver, heart, and brain tissue homogenates prepared with 6% perchloric acid.[19] The NAD+/NADH ratios were calculated according to Krebs. Methemoglobin in the hemolysate of erythrocytes was determined as per the method of Winterbourn.[20] Proteins were determined according to the Lowry method. DPPH assay Scavenging of DPPH free radicals was measured as described elsewhere.[21] Briefly, splenozide (stock solution) and GSH (0.1 mM final concentration) were mixed with 0.1 mM DPPH solution in a cuvette (all dissolved in ethanol). The time course of the optical density change was determined at 520 nm at 25°C. We defined the period of DPPH semi-transformation (T 50 ) into the nonradical form and the velocity constants for these reactions (K) under the influence of splenozide and GSH. Statistical analysis Data were expressed as mean±SEM and the differences among groups were analyzed using one-way analysis of variance (ANOVA) followed by Bonferroni's multiple comparison test. P< 0.05 was considered significant. Results Nitrite intoxication Acute nitrite intoxication induced the increase of methemoglobin (MetHb) up to 34% in rat blood. Nitrite in toxic dose caused a significant (P < 0.001) increase in MDA content in the liver, heart and brain by 56%, 42%, 86%, respectively, compared to control. [Figure - 1] Simultaneously in these tissues, there was a decrease in the activities of superoxide dismutase (SOD), CAT, GPx, GR, and a reduction in GSH content. [Table - 1] In the heart, the decrease in CAT activity was insignificant. In all the tissues there was enhanced cytosolic G6PDH activity and a decrease in mitochondrial SDH activity. Nitrite poisoning caused an increase in lactate concentration and diminution of NAD+/NADH rate. [Table - 2] Nucleosides complex and nitrite intoxication After the splenozide pretreatment, the tendency for MetHb-content lowering was registered in blood (from 34% to 29%). Splenozide injection induced a reduction of LPO process in tissues. [Figure - 1] In the liver, heart and brain, MDA concentration decreased by 19%, 23% and 18%, respectively, compared to acute intoxication ( P < 0.001). In the heart, SOD and CAT activities increased by 32% ( P < 0.001) and 14% ( P < 0.01), respectively; although in the liver and brain the increase was not significant. A significant ( P < 0.001) increase in G6PDH activity was seen in liver (35%), myocardium (51%), and brain (21%). The GSH content and the activities of GR and GPx increased in all experimental tissues. [Table - 1] Splenozide decreased lactate and increased NAD+ forms. The SDH activity was enhanced in the heart while in liver and brain the changes were not statistically significant. [Table - 2] Nucleosides and DPPH radical scavenging As shown in [Figure - 2], under the splenozide influence the period of DPPH semi-transformation into the nonradical form is T 50 = 2 days, velocity constant for this reaction is K= 1,01 l/molFNx01min, and these values are lower than the same ones for GSH (K = 41602 l/molFNx01min, T 50 =10s) . Discussion Nitrite methemoglobinemia is a potent process for free-radical generation.[3] It is known that nitrite can stimulate oxidation of ferrous ions in oxyhemoglobin to form MetHb and superoxide-anion radicals.[2],[3] This xenobiotic is a ready source of nitric oxide which reacts rapidly with superoxides to form highly reactive peroxinitrite (ONOO-).[2] Reactive oxygen metabolites can initiate a wide range of toxic oxidative reactions. These processes include initiation of LPO, oxidative modification of proteins, direct inhibition of mitochondrial respiratory chain enzymes, inactivation of membrane Na+/K+ ATP-ase activity, and disorders in the antioxidant defense system.[22] In the present study, acute nitrite intoxication caused a significant increase in MDA content, a decrease in the activities of SOD and CAT, and a depletion of the intracellular GSH pool. Depression of the GR-function activity can evoke changes in the ratio of oxidized and reduced glutathione forms, with accumulation of oxidized equivalents which are oxidative stress markers. [Table - 1] Reduction of GPx activity in NaNO 2 -treated rats [Table - 1] may be due to enhanced ONOO- production. It is known that ONOO- reacts nonenzymatically with selenium in the active site of GPx , resulting in a further inactivation of this enzyme.[6] The elevated levels of MetHb (~34%) in the present study could have been the cause of anemic hypoxia, due to an inadequate supply of oxygen to the tissues. Our experiments with mitochondrion fraction demonstrated that nitrite injection inhibited SDH activity, the key enzyme of electron transport in mitochondria (Complex II), which was followed by oxidative phosphorylation disconnection and energy limitation.[23] One of the most important consequences of energy deficiency under hypoxia is altered redox state of the cell. We registered a decrease in NAD+/NADH rate and cytosolic pH in the tissues intoxicated with NaNO[2] and this is an evidence of intracellular acidosis development. These changes can have an influence on other redox-dependent reactions, i.e. oxidation of ferrous ions, promoters of ROS generation (through the Fenton reaction), peroxidation of lipids, and antioxidant status of the tissues.[22] Splenozide pretreatment decreased LPO process in all the studied tissues, while its effect on antioxidative enzymes (SOD, CAT) depended on the tissue type. Splenozide significantly increased G6PDH activity, an enzyme that catalyzes the initial rate-limiting step in the pentose phosphate pathway and is responsible for the generation of NADPH.[24] It is possible that in case of complex nucleosides pretreatment, the level of intracellular GSH was maintained by the GSH recycle activation via., GR in NADPH-dependent reactions. Glutathione, through its antiradical, antioxidant, signal function and maintenance of redox- balance properties, plays a vital role in cell function. Glutathione, the main storage form of sulfur, acts as a potent detoxifier of xenobiotics through GSH conjugation.[25] Our in vitro study demonstrated a weak antiradical property of splenozide that could be exerted by the optimization of the glutathione system. Splenozide pretreatment in NaNO 2 poisoning led to the decrease in lactate concentration, accumulation of oxidized NAD and decline in metabolic acidosis. It is possible that splenozide, being a nucleoside complex, quickly penetrates into the tissues.[8],[9] This complex influences on the redox status and as a consequence, on cell metabolism. It was reported that exogenous nucleosides and their analogues influence nucleic acid synthesis, coenzyme de novo , and intensify processes in tissues.[9] According to the present study, myocardial tissue was the most sensitive to splenozide action. This effect not only strengthened antioxidant defense, but also quickly restored the electron transport function of the mitochondrial respiratory chain. It may be concluded that splenozide pretreatment decreases the tissue oxidative stress in acute nitrite intoxication by maintaining the GSH recycling activity through activation of NADPH-dependent reactions, as well as through its antiacidotic action. Acknowledgments The authors are grateful to Dr. Olyinik of Komissarenko Institute of Endocrinology and Metabolism, National Academy of Sciences (Kiev, Ukraine) for the supply of splenozide and the suggestions towards the study.References
Copyright 2006 - Indian Journal of Pharmacology The following images related to this document are available:Photo images[ph06117t2.jpg] [ph06117t1.jpg] [ph06117f1.jpg] [ph06117f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}