 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
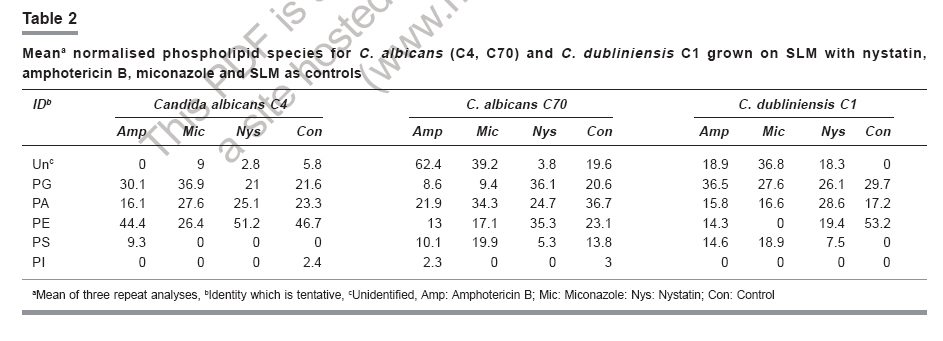
Indian Journal of Pharmacology, Vol. 38, No. 6, November-December, 2006, pp. 423-426 Research Paper Effect of amphotericin B, nystatin and miconazole on the polar lipids of Candida albicans and Candida dubliniensis Mahmoudabadi AZ, Drucker DB Department of Medical Mycoparasitology, Jundishapour University of Medical Sciences, Ahwaz Date of Submission: 29-Dec-2005 Code Number: ph06119 Abstract Objective : To determine whether nystatin, amphotericin B and miconazole have similar effects upon the fatty acids and phospholipids of Candida albicans and C. dubliniensis .Materials and Methods: Serial dilutions of antifungal drugs were prepared in flasks. Then 50 ml of standardised suspension of tested yeasts was inoculated into each flask and incubated in shaking water bath at 37°C for 48 h. The last flask that had growth was centrifuged and the yeast cells harvested, washed and freeze-dried. Polar lipids were extracted from freeze-dried cells and were analysed by fast atom bombardment mass spectrometry (FAB MS) in negative-ion mode. Results: Nystatin, amphotericin B and miconazole have different effects on phospholipids and fatty acids of two strains of C. albicans and a single strain of C. dubliniensis . The content of phosphatidylethanolamine (PE) in C. dubliniensis decreased from 53.2% to 19.4% and from 53.2% to 14.3% in the presence of nystatin and amphotericin B, respectively, whereas this phospholipid was absent in cultures exposed to miconazole. In both the examined strains of C. albicans , PE was decreased,in the presence of amphotericin B and nystatin, whereas PE in both strains of C. albicans increased when cultures were exposed to nystatin. Conclusion: It is concluded that biosynthesis of fatty acids and phospholipids of C. albicans and C. dubliniensis is affected by nystatin, amphotericin B and miconazole, in addition to the effects on ergosterol previously described. Antifungals also exert both qualitative and quantitative effects on different strains of C. albicans. Keywords: Antifungal antibiotics, azoles, candidiasis, cell membrane. Introduction Candida albicans is saprophytic yeast that causes candidiasis. Superficial candidiasis is usually treated with a topical preparation such as nystatin or miconazole, while disseminated infections require systemic therapy, e.g., amphotericin B. Amphotericin B, nystatin and miconazole inhibit the growth of many of fungi in vitro . The antifungal activity of these drugs has been reported to be due to an interaction with sterol in the cell membrane of fungus, especially the ergosterol.[1] The polyene-resistant mutants of C. albicans have a different phospholipid fatty acyl composition.[2] The differing fatty acid composition has been suggested as a reason for the variation in polyene sensitivity to C. albicans .[3] Azoles interact directly with the membrane lipids without necessarily binding to them.[4],[5] For example, interaction with cell membrane phospholipids and fatty acids causes leakage of proteins and amino acids.[6],[7] Miconazole interacts with phospholipids and changes the lipid organisation without binding to the lipids.[7],[8] Some of the antifungal agents can inhibit fatty acid synthesis in yeasts.[8],[9],[10] For example, cerulenin is a specific inhibitor of fatty acid.[11] However it has also been shown that fatty acids are capable of protecting Saccharomyces cerevisiae against the action of polyenes.[12] Palmitoleic acid (C 16:1 ), oleic acid (C 18:1 ), α-linoleic acid (C 18:2 ), γ-linoleic acid (C 18:3 ) and arachidonic acid (C 20:4 ) have been reported to possess an antagonistic effect against clotrimazole and miconazole.[13] Moreover, the fatty acid content of the polyene sensitive cultures of C. glabrata differs from the polyene- resistant cultures.[14] The imidazoles, such as miconazole, econazole, clotrimazole and ketoconazole, decrease the unsaturated-saturated ratio of fatty acids in vivo .[15] Chrysodin has been reported to significantly decrease the percentage composition of linolenic acid and linoleic acid, and to increase that of oleic acid.[10] The linoleate content in the phospholipid fraction of C. albicans increased from 15% to 25% when incubated in the presence of ketoconazole or miconazole.[8] The contents of phosphatidylinositol (PI), phosphatidylethanolamine (PE) and phosphatidylcholine (PC) decrease with validamycin A.[16] The aim of this study was to determine whether amphotericin B, nystatin and miconazole have similar effects upon the fatty acids and phospholipids of C. albicans and C. dubliniensis . Materials and Methods Organisms and preparation of the inoculum Candida albicans and C. dubliniensis were obtained from the patients of Dental Hospital, the University of Manchester. Isolates were confirmed as C. albicans (C4, C70) and C. dubliniensis (C1) by production of yellow-green to blue-green colonies on CHROMagar Candida, germ tube tests on horse serum, and by production of chlamydoconidia on cornmeal agar, growth at 45°C and ID 32 C kits. Isolates were maintained on Sabouraud's Dextrose Agar (SDA) (BBL, Becton Dickinson, USA). Synchronous cultures were prepared according to the method of Johnson. [17] Briefly, yeast isolates were sub-cultured on SDA and incubated at 37°C for 24 h. Yeast cells were collected in 2 ml of sterile phosphate buffer solution (PBS) with sterile cotton wool swabs to prepare a suspension. The suspension was adjusted to 70% T by a spectrophotometer (Pye Unicam) set at 530 nm, which was expected to result in a suspension containing about 1 X 10 6sub cfu/ml. Test method Sixty-four mg of amphotericin B (Sigma, Poole, Dorset, UK), nystatin (Sigma) and miconazole (Sigma) were separately dissolved in 50 ml of dimethyl sulphoxide (Sigma) at 1280 mg/l. The solutions were left at room temperature for 30 min for self-sterilisation and then stored at -70°C for up to 1 month. The stock solutions of amphotericin B and nystatin were diluted to 640 mg/l with sterile distilled water. Serial dilutions of antifungals (nystatin and amphotericin B, 32-0.125 mg/l; miconazole, 64-0.25 mg/l) were prepared in flasks containing Sabouraud liquid medium (SLM) (Oxoid, Hampshire, England) and inoculated with 50 µl of standardised suspension of tested yeasts. The flasks were then incubated in shaking water bath at 37°C for 48 h. The last flask that had growth was centrifuged and the yeast cells harvested, washed and freeze dried. Lipid extraction and analysis Polar lipids were extracted from freeze-dried cells with chloroform-methanol (1:2, v/v) and analysed by fast atom bombardment mass spectrometry (FAB MS) in negative-ion mode with a Concept IS mass spectrometer (Kratos, Manchester, UK). All experiments were repeated three times. Fatty acids were presented as low mass ions ( m/z 200-300), while phospholipid analogue anions were seen as high mass anions ( m/z 500-1000). The 10 most abundant anion peaks per isolate observed in the mass spectra were selected for data analysis. Results were entered into a spreadsheet (Excel) and normalised (S% peaks = 100). Results Candida albicans Nystatin: The minimal inhibitory concentration (MIC) of nystatin for both isolates of C. albicans was 0.25 mg/l; the polar lipids data were obtained from cells growing on the next serial dilution (0.125 mg/l). Carboxylate anion profiles of both tested Candida grown on nystatin showed that C 18:1 was the major fatty acid in spectra. When C. albicans grew on SLM + nystatin the biosynthesis of m/z 297(C 19:0 ) in C. albicans (C70) and 227(C 14:0 ) in C. albicans (C4) were inhibited, whereas the contents of m/z 281(C 18:1 ) and 279(C 18:2 ) were increased in both isolates. [Table - 1] The content of phosphatidic acid (PA) in C. albicans (C70) was proportionately decreased compared to control from 36.7% to 24.7%. [Table - 2] Other phospholipids, such as PE and phosphatidylglycerol (PG), were considerably increased from 23.1% to 35.3% and 20.6% to 36.1%, respectively. Amphotericin B: The MIC of amphotericin B for both isolates of C. albicans was 0.25 mg/l and the polar lipids of these strains were obtained from cultures grown on the next serial dilution of drug (0.125 mg/l). While amphotericin B decreased the content of m/z 253(C 16:1 ), 255(C 16:0 ), 279(C 18:2 ), 281(C 18:1 ) from a combined amount of 80.9% of total acid anions in control to only 41.3%, on the other hand, 283(C 18:0 ) was increased from 1.4% in controls to 19.9%. Amphotericin B inhibited the biosynthesis of m/z 239(C 15:1 ), 241(C 15:0 ), 267(C 17:1 ) and 297(C 19:0 ) in C. albicans (C70) and m/z 297(C 19:0 ) and 267(C 17:1 ) in C. albicans (C4). [Table - 1] PA and PE were decreased in both C. albicans isolates, whereas PG decreased from 20.6% in control to 8.6% in C. albicans (C70) and increased in C. albicans (C4). [Table - 2] Miconazole: The MIC of miconazole for both tested C. albicans (C4, C70) was 2 mg/l and polar lipids were obtained from growing on the next serial concentration (1 mg/l). The carboxylate anions detected after-growth of C. albicans (C70) with miconazole induced m/z 221(C 14:3 ) and 269(C 17:0 ), which were not detected in controls. Miconazole, also, inhibited the biosynthesis of m/z 267(C 17:1 ) in C. albicans (C70) and decreased the amount of m/z 253(C 16:1 ) and 279(C 18:2 ). [Table - 1] The content of PA, PG and PE were decreased compared with C. albicans (C70). The content of PA, PG and PE were considerably increased in C. albicans (C4). [Table - 2] Candida dubliniensis Nystatin: The MIC of nystatin for C. dubliniensis (C1) was 1 mg/l, and the polar lipids were obtained from growing on the next serial dilution (0.5 mg/l). Nystatin caused the proportion of m/z 253(C 16:1 ), 255(C 16:0 ), 279(C 18:2 ) and 281(C 18:1 ) to decrease, while the proportions of m/z 283(C 18:0 ) and 241(C 15:0 ) increased. The proportion of 295(C 19:1 ) was only seen in C. dubliniensis (C1) when exposed to nystatin. [Table - 1] The content of PE and PG generally decreased in the presence of nystatin whereas PA increased. Phosphatidylserine (PS) was found in C. dubliniensis when exposed to nystatin whereas controls had none detectable. [Table - 2] Amphotericin B: In this study, the MIC of amphotericin B for C. dubliniensis (C1) was 1.0 mg/l and the polar lipids were obtained from growing on the next serial dilution of amphotericin B (0.5 mg/l). [Table - 1] shows that m/z 255(C 16:0 ) (25.4%) was the major carboxylate anion in culture growth with amphotericin B compared to m/z 281(C 18:1 ) (28.1%) in control. When C. dubliniensis grew on amphotericin B, the greatest variation in carboxylate anion was observed in the content of m/z 295(C 19:1 ), 281(C 18:1 ) and 279(C 18:2 ). The proportion of PE decreased considerably from 53.2% in controls to 14.3% in cultures exposed to amphotericin B. The next most intense phospholipid families were PA (15.8%) and PE (14.3%). [Table - 2] Miconazole: The MIC of miconazole for C. dubliniensis was 2.0 mg/l and the polar lipids were obtained from cells grown on the next serial dilution of drug (1.0 mg/l). Miconazole decreased the proportions of m/z 253 (C 16:1 ) and 281 (C 18:1 ) from 19.4% to 13.8% and 28.1% to 12.2%, respectively. Miconazole completely inhibited the biosynthesis of m/z 279(C 18:2 ) and 277(C 18:3 ). [Table - 1] PA and PG proportions were nearly similar in cultures with miconazole or control, whereas PE was absent. [Table - 2] Discussion Many researchers have studied the mechanisms by which amphotericin B, miconazole and nystatin affect both the morphology of C. albicans cells and their mechanisms of drug resistance. However, the effect of antifungals on phospholipid analogues and fatty acids of yeasts has not been examined. Our results showed that the content of polar lipids (carboxylic acids and phospholipids) was changed in Candida isolates when grown with nystatin, miconazole and amphotericin B. Koul et al. , [18] have suggested that lipids are targeted by some of the anti Candida drugs. Van Den Bossche et al. ,[8] have suggested that this shift, to more unsaturated fatty acid, results in increased membrane fluidity. According to Niimi et al. ,[19] unsaturated fatty acids bind to azole (clotrimazole). Hitchcock et al. ,[2] have found that changes in phospholipid composition affect polyene sensitivity. In this study nystatin caused the ratio of unsaturated-saturated fatty acids to increase in both species,compared with controls. Nystatin causes abnormal growth of C. albicans .[20] Iannitelli and Ikawa[12] have shown that fatty acids decrease the activity of nystatin against S. cerevisiae . Also, longer chain-length fatty acids, and those more unsaturating, are more effective. Nystatin had different effects on phospholipids of tested yeasts. For example, the content of PE and PG were increased in C. albicans C70 but proportionately decreased both phospholipids in C. dubliniensis C1. Azoles have a direct effect on the fatty acids of cell membranes.[6] Georgopapadakou et al. ,[15] have reported that miconazole increases the content of C 16:0 and C 18:0 whereas it decreases the proportion of C 18:2 and C 18:3 in vitro . In our study the proportion of C 18:0 increased in both isolates of C. albicans when exposed to miconazole, whereas C 18:2 increased proportionately in one strain but decreased in the other. Long-chain fatty acids, palmitoleic acid (C 16:1 ), oleic acid (C 18:1 ), α-linoleic acid (C 18:2 ), γ-linoleic acid (C 18:3 ) and arachidonic acid (C 20:4 ) have antagonistic effects against miconazole.[13] The content of linoleate (C 18:2 ) was found to be 15% of the total fatty acid in phospholipid fractions of C. albicans , while that increased up to 25% in the phospholipid fractions of C. albicans grown in the presence of miconazole.[8] The presence of miconazole mainly affects the proportions of C 18:1 and C 16:1 in spectra. This possibly indicates that fatty acids are involved in the binding site of the drug at the membrane. Georgopapadakou et al. ,[15] have reported that miconazole has no effect on C 18:1 . However in our study, the proportion of C 18:1 was considerably decreased in all tested yeasts. In our study PA, PG and PE proportions decreased in one isolate of C. albicans and increased in the other isolate. The proportions of phospholipid classes in C. dubliniensis exposed to miconazole were similar to data for controls (no miconazole). Amphotericin B had the most effect on C 18:0 and increased the proportion of C 18:0 14.2 times in C. albicans (C70) , as compared to control. The major fatty acid in control was C 18:1 compared to C 18:0 in amphotericin B cultures. In C. dubliniensis , the greatest changes were seen with the proportion of C 18:2 and C 19:1 . In this species the ratio of unsaturated to saturated fatty acids decreased. Generally, amphotericin B caused PS synthesis in C. dubliniensis and one isolate of C. albicans . Also, PA and PE proportions decreased in both C. albicans strains, whereas PG increased in C. albicans C4 only. The proportion of unsaturated fatty acids in both species increased when exposed with nystatin, whereas amphotericin B and miconazole decreased the proportion of unsaturated fatty acids. Conclusion It is concluded that the biosynthesis of fatty acids and phospholipids are affected by nystatin, amphotericin B and miconazole, in addition to effects on ergosterol as previously described. Also antifungals have different effects on polar lipids of different strains of C. albicans. References
Copyright 2006 - Indian Journal of Pharmacology The following images related to this document are available:Photo images[ph06119t1.jpg] [ph06119t2.jpg] |
| |||||||||

{kind=link}
{kind=link}