 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
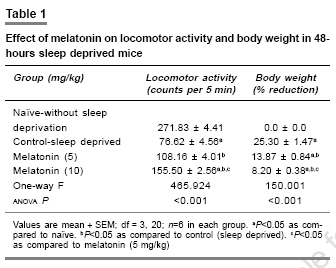
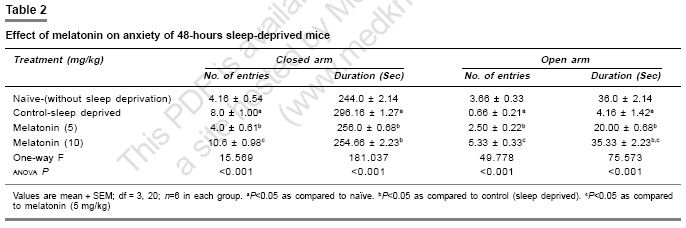
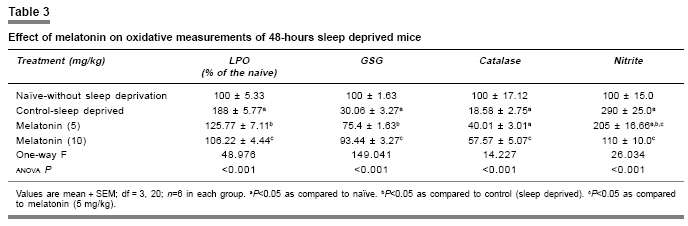
Indian Journal of Pharmacology, Vol. 39, No. 1, January-February, 2007, pp. 48-51 Research Paper Protective effect of melatonin on certain behavioral and biochemical alterations induced by sleep-deprivation in mice Kalonia Harikesh, Kumar Anil Pharmacology Division, University Institute of Pharmaceutical Science, Panjab University, Chandigarh- 160014 Date of Submission: 29-May-2006 Code Number: ph07011 Abstract Objective: To explore the effect of melatonin on the behavioral and biochemical parameters in sleep-disturbed mice.
Keywords: Anti-oxidant, insomnia, pineal hormone Introduction Sleep is normally a period of relaxation and repair, important for the maintenance of physiological homeostasis and psychological balance. The disturbance or shortening of normal sleep has recently been reported to produce harmful effects to metabolic and endocrine functions.[1] Sleep deprivation causes decrease in body weight despite an increase in food intake,[2] initial hyperthermia followed by hypothermia,[3] increase in anxiety levels[4] and decrease in locomotor activity, irritability,[5] and poor performance.[6] Sleep deprivation is also known to impair cognitive functions.[7] Sleep deprivation results in an increase in plasma glucocorticoids in both human and rodents.[8] Moreover, sleep deprivation also leads to augmentation of the aging process because of increased brain levels of cortisol which cause a gradual damage to brain cells.[9] Sleep deprivation has also been proposed to cause oxidative stress.[10] Brain is more susceptible to peroxidation because it is one of the most metabolically active parts of the body. Furthermore, neural tissue is rich in polyunsaturated fatty acids and its antioxidant status is lower than that of other tissues, i.e., it has low levels of catalase activity.[11] Melatonin (N-acetyl-5-methoxy-tryptamine), a hormone of the pineal gland[12] has several important functions, such as circadian rhythm regulation, immune enhancement, sleep induction, seasonal reproductive regulation, and light-dark signal transduction. Melatonin is also an effective free radical scavenger and antioxidant.[13] It has been shown to be highly useful in preventing neural degenerative changes.[14] Several studies report that melatonin has mild to moderate hypnotic action in animals.[15] However, the effect of melatonin in sleep-disturbed mice is yet to be established. Therefore in the present study, the protective effects of melatonin in sleep-disturbed mice were investigated. Materials and Methods Animals Inbred, male laca mice (25-30 gram) were maintained under standard laboratory conditions of 12-hour light/dark cycle with free access to food and water . Animals were acclimatized to laboratory conditions before the test. Each animal was used once in the experiments. All the experiments were performed between 9:00 and 17:00 hours. The experimental protocol was approved by the Institutional Animal Ethics Committee and was conducted according to the Indian National Science Academy Guidelines for the use and care of experimental animal, drug, dose, and treatment schedule. The animals were divided into six groups, having six to nine animals each group. The first and second groups were treated as naοve (without sleep deprivation) and 48 hours sleep deprived (suspended on grid), respectively. Groups three and four were injected with 5 and 10 mg/kg melatonin (dissolved in 0.1% dimethylsulfoxide [DMSO]), respectively. All the drugs were administered in morning session (9:00-10:00 AM) intra-peritoneally, once a day for five days, started three days before 48 hours-sleep deprivation. Sleep deprivation Animals were sleep deprived by placing on a grid suspended over water method developed by Shinomiya.[16] In this method, animals were placed on a grid floor (29´15´7 cm) inside the plastic cage filled with water to 1 cm below the grid surface for 48 hours. The stainless steel rods of the grid (3 mm wide) were set 2 cm apart from each other. Food and water were provided ad libitum. Body weight change The body weight of animals was recorded before the start of experiment and thereafter before behavioral quantification. Measurement of anxiety levels Elevated plus maze, described by Kulkarni[17] is a novel test to study the anxiogenic and anxiolytic drug effects in rodents. The plus maze apparatus consist of two open (16´5 cm [for mice]) and two closed arms (16´5´12 cm) and is placed at a height of 25 cm (for mice). The animals were placed individually at the center of the maze with the head facing the open arm. During the 5 min test, the number of entries into the open and closed arms and the time spent in each arm were recorded. Locomotor activity The locomotor activity was monitored using actophotometer (IMCORP, India). The animals were individually placed in activity meter for three minutes before counting of actual locomotor activity for the next 5 minutes. The locomotor activity was expressed in terms of total photobeams counts/5 min per animal.[18] Dissection and homogenization On day 5, the animals were sacrificed by decapitation immediately after behavioral assessments. The whole brains were removed, tissue homogenate (10% w/v -1) was prepared in 0.1 m phosphate buffer (pH 7.4), centrifuged for 20 minutes at 15000 r.p.m. and the supernatant estimated for of lipid peroxidation and reduced glutathione levels. The post nuclear fractions for catalase assay were obtained by centrifugation of the homogenate at 1000 g for 20 min, at 4°C and for other enzyme assays centrifuged at 12,000 g for 60 min at 4°C. Lipid peroxidation assay The quantitative measurement of lipid peroxidation was performed according to the method of Wills.[19] The amount of malondialdehyde, a measure of lipid peroxidation was estimated by reaction with thiobarbituric acid at 532 nm using a lambda 20 spectrophotometer (Perkin Elmer). The values were calculated using molar extinction coefficient of chromophore (1.56´10 5sub M -1 cm -1) and expressed as percentage of control. Estimation of reduced glutathione Reduced glutathione in brain was estimated according to the method of Ellman.[20] About 1 mL supernatant was precipitated with 1 mL of 4% sulfosalicylic acid and cold digested at 4°C for 1 hour. The sample was centrifuged at 1200 g for 15 min at 4°C. To 1 mL of this supernatant, 2.7 mL of phosphate buffer (0.1m, pH 8) and 0.2 mL of 5,5-dithiobis (2-nitrobenzoic acid [DTNB]) were added. The yellow color developed was read immediately at 412 nm using a spectrophotometer (Shimadzu). Results were calculated using molar extinction coefficient of chromophore (1.36´10 4 m -1cm -1) and expressed as percentage of control. Estimation of nitrite The accumulation of nitrite in the supernatant, an indicator of the production of nitric oxide (NO), was determined with a colorimetric assay with Greiss reagent (0.1% N- (1-Napthyl) ethylenediamine dihyrochloride, 1% sulfanilamide and 2.5% phosphoric acid) as per the method of Green et al .[21] Equal volumes of supernatant and Greiss reagent were mixed, incubated for 10 min at room temperature and the absorbance read at 540 nm using a spectrophotometer. The concentration of nitrite in the supernatant was determined from a standard curve and expressed as percentage of control. Protein estimation The protein content was measured according to the method of Lowry[22] using bovine serum albumin as standard. Estimation of catalase Catalase activity was assayed by method of Luck.[23] Briefly, the assay mixture consisted of 3 mL of H 2 O 2 phosphate buffer (1.25´10 -2 M H 2 O 2 ) and 0.05 mL of supernatant of the brain homogenate (10%) and the change in absorbance was read at 240 nm using the spectrophotometer. Enzyme activity was calculated using the millimolar extinction coefficient of H 2 O 2 (0.07). The results were expressed as micromoles of H 2 O 2 decomposed per min, per mg of protein. Statistical analysis One specific group of mice was assigned to a specific drug treatment each group consist of six to nine ( n =6-9) animals. All the values are expressed as mean ± SEM. The data were analyzed by analysis of variance (anova) followed by Tukey test. The criterion for statistical significance was P < 0.05. Results Melatonin effect on body weight and locomotor activity Body weights and locomotor activity of 48-hours sleep-deprived mice were significantly reduced as compared to naοve mice (without sleep deprivation). Melatonin pretreatment (5 and 10 mg/kg) for 5 days, significantly reversed the reduction in body weight and locomotor activity ( P < 0.05) [Table - 1]. Effect of melatonin on anxiety level About 48-hours sleep deprivation caused severe anxiety in mice (increased the number of entries and duration in closed arm and decreased the number of entries as well as duration in open arm of plus maze performance task), which was significant as compared to naοve mice. Pre-treatment with melatonin (5 and 10 mg/kg) increased the number of entries and duration of time spent in open arm as well as decreased the number of entries and duration in closed arm significantly ( P < 0.05) as compared to control (sleep deprived) [Table - 2]. Glutathione and catalase activity About 48-hours sleep deprivation decreased glutathione level and catalase activity significantly as compared to naοve (without sleep deprivation) mice. Pretreatment with melatonin (5 and 10 mg/kg) restored significantly ( P < 0.05) the depleted glutathione and catalase activity in sleep deprived animals [Table - 3]. Lipid peroxidation and nitrite level About 48-hours sleep loss significantly increased lipid peroxidation and the nitrite level, compared to naοve (without sleep deprivation) P < 0.05. Melatonin pretreatment (5 and 10 mg/kg) significantly ( P < 0.05) reversed malondialdehyde and NO activity as compared to the control (sleep deprived) [Table - 3]. Discussion Sleep deprivation is considered a risk for development of certain diseases. The behavioral and biochemical changes produced by sleep deprivation that results in health consequences, however, are largely unknown. Sleep deprivation caused various behavioral disturbances involving motor activity, anxiety level, memory and metabolic functions related to anabolic hormones, body weight, and so on.[4] Sleep deprivation has recently been reported to cause oxidative stress. Ramanathan et al.[10] also reported a significant decrease in superoxide dismutase activity in the hippocampus and brain stem in sleep-deprived rats. Present model of sleep disturbance is also used to evaluate hypnotic drugs. In the present study, 48-hours sleep deprivation in mice significantly reduced locomotor activity, body weight and increased anxiety and melatonin pretreatment (for 5 days) significantly prevented these changes. Marked behavioral changes could be hypothesized due to pathological changes in the brain regions involved in sleep regulation. Sleep deprivation has been reported to cause hypermetabolism.[1] In addition, 48-hours sleep deprivation also increased lipid peroxidation and nitrite, accompanied by a decrease in glutathione and catalase activity, suggesting oxidative stress in sleep deprived animals. However, little is known whether stress is an important consequence of sleep deprivation. Reimund theorized[24] that sleep increases the efficiency of antioxidant mechanisms in the brain. In present study, melatonin significantly reduced the lipid peroxidation, nitrite level and restored catalase activity and glutathione level in mice brain. Melatonin is not only a non-enzymatic scavenger, but also an activator of antioxidative enzymes such as superoxide dismutase, glutathione peroxidase.[25] From the present study it is tempting to suggest that the protective effect of melatonin could be due to its antioxidative properties. Conclusion Present study suggests that melatonin has protective action against sleep deprivation-induced behavioral and biochemical alterations and can be considered in the management of sleep related problems.References
Copyright 2007 - Indian Journal of Pharmacology The following images related to this document are available:Photo images[ph07011t1.jpg] [ph07011t2.jpg] [ph07011t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}