 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
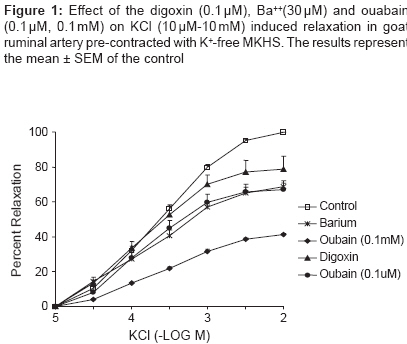
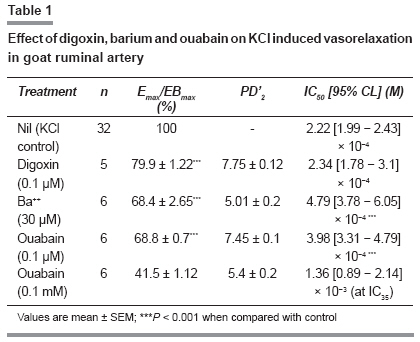
Indian Journal of Pharmacology, Vol. 41, No. 2, March-April, 2009, pp. 68-71 Research Article Role of Na + -K + ATPase enzyme in vascular response of goat ruminal artery Kathirvel K, Parija SC Department of Pharmacology and Toxicology, Faculty of Veterinary Sciences and Animal Husbandry, Orissa University of Agriculture and Technology, Bhubaneswar - 751 003 Date of Submission: 21-Jan-2008 Code Number: ph09020 DOI: 10.4103/0253-7613.51343 Abstract Objective: To study the role of Na + , K + - ATPase enzyme in the vascular response of goat ruminal artery.Materials and Methods: Ruminal artery was obtained in chilled aerated modified Krebs-Henseleit solution (KHS) from a local slaughterhouse and transported in ice for further processing. The endothelium intact arterial ring was mounted in a thermostatically controlled (37 ± 0.5°C) organ bath containing 20 ml of modified KHS (pH 7.4) bubbled with oxygen (95%) and CO 2 (5%) under 2g tension. An equilibration of 90 min was allowed before addition of drugs into the bath. The responses were recorded isometrically in an automatic organ bath connected to PowerLab data acquisition system. In order to examine intact functional endothelium, ACh (10µM) was added on the 5-HT (1.0µM) - induced sustained contractile response. Similarly, functional characterization of Na + , K + -ATPase activity was done by K + -induced relaxation (10µM-10mM) in the absence and presence of ouabain (0.1µM/ 0.1mM), digoxin (0.1µM) and barium (30µM). Results: ACh (10-5 M) did not produce any relaxing effect on 5-HT-induced sustained contractile response suggesting that vascular endothelium has no significant influence on the activation of sodium pump by extracellular K + in ruminal artery. Low concentration of Ba 2+ (30 µM) (IC 50 : 0.479mM) inhibited K + -induced relaxation suggesting K ir (inward rectifier) channel in part had role in K + -induced vasodilatation in ruminal artery. Vasorelaxant effect of KCl (10µM-10mM) in K + -free medium is also blocked by ouabain (0.1µM and 0.1mM) (IC 50 :0.398mM and IC 35 : 1.36mM), but not by digoxin (0.1µM) (IC 50 0.234mM) suggesting that ouabain sensitive Na + , K + -ATPase isoform is present in the ruminal artery. Conclusion: In the goat ruminal artery functional regulation of sodium pump is partly mediated by K + channel and ouabain sensitive Na + , K + ATPase. Keywords: Barium, K + channel, K + - ATPase, Na + , oubain, ruminal artery Introduction Na + ,K + -ATPase is an enzyme of the plasma membrane of most cells that uses cellular ATP to exchange cytoplasmic Na + for extracellular K + . [1] The function of the Na,K-ATPase is essential for the generation and maintenance of the electrochemical gradients. [2] Structurally, Na + ,K + -ATPase is an oligomer that is composed of distinct molecular forms of two major polypeptides, the α and β subunits. [3] At present, four structural variants of the α polypeptide (α1 , α2 , α3 and α4 ) and three b (b1 , b2 and b3 ) subunits have been identified in mammals. Association of the a and β polypeptides in different oligomers results in multiple isozymes of the Na + ,K + -ATPase that have unique functional properties and a tissue-specific pattern of expression. [3] The sodium pump, in turn, is the target for multiple regulatory mechanisms. [4] It is also responsible for maintaining tone and contractility of smooth muscle. [5] Ouabain, a cardiotonic steroid, has been shown to be an endogenous factor that is secreted by the adrenal glands in humans and other mammals and is present in blood at nanomolar concentrations. [6] The mechanism of action of ouabain has been attributed classically to ion changes that are secondary to inhibition of the catalytic and transport activity of the Na + ,K + -ATPase. [7] Four different isoforms, namely α1 , α2 , α3 and α4 with different ouabain affinities are expressed in different species and in a tissue-specific manner. [8] Regional variations in activity and isoform-specific expression of sodium pump in vascular tissues have been reported. [9] The presence of a microsomal Na + ,K + -ATPase in sheep rumen epithelium activity which is reduced by 50% in the presence of ouabain has been reported. [10] Studies using biopsies of rumen epithelium papillae measured a net influx of [ 86 Rb] across the canine ruminal epithelium [11] and these findings are similar to a high concentration of Na + , K + -ATPase found in [ 3 H] ouabain-binding studies. [12] But there is no research related to ruminal artery to date on Na + , K + -ATPase and its role in maintaining the vascular tone in ruminal artery. Therefore, the present study was undertaken to identify the sensitivity of Na + , K + -ATPase to ouabain in goat ruminal artery. Materials and Methods Preparation of ruminal arterial ring and tension recording ACh (10 -6 M) on 5-HT- induced sustained contractile response Experiments with K+-free physiological solution KCl-induced relaxation in K + - free MKHS in presence of barium KCl-induced relaxation in K + - free MKHS in presence of digoxin KCl-induced relaxation in K + - free MKHS in presence of ouabain Data analysis Drugs Results Effect of ACh on 5-HT (0.1µM) induced contraction and KCl on K+-free medium Effect of digoxin, barium and ouabain on KCl-induced relaxation The mean maximal inhibition (I Bmax ) of KCl-induced relaxation response curve was significantly ( P < 0.001) by Ba 2+ (31.7 ± 2.7%), digoxin (20.1 ± 1.2%), ouabain at 0.1µM (31.2 ± 0.7%) and 0.1mM (58.5 ± 1.2%) as compared to pooled control (100%). The KCl -induced relaxation response curve was shifted to right in presence of Ba 2+ , ouabain at 0.1µM and 0.1mM with significant decrease in the mean IC 50 . In the presence of ouabain (0.1mM), there was clear-cut rightward shift of the KCl-induced inhibitory response curve without attainment of IC 50 . As in the presence of ouabain (0.1mM) the maximal inhibition was less than the 50% (E Bmax : 41.5 ± 1.12%), the IC 35 was calculated for comparison. The IC 35 of the KCl-induced inhibitory response curve in presence of ouabain (0.1mM) (IC 35 :1.36mM) was about 10.23 fold more than the control (IC 35 : 0.139mM). The mean I Bmax , pD ′ 2 and IC 50 of the KCl-induced relaxation response curve in presence of Ba 2+ or digoxin or ouabain were compared with the control in the [Table - 1] and curves are presented in [Figure - 1]. Discussion Na + -K + -ATPase exists in the plasma membrane as a heterodimer consisting of a catalytic α-subunit and a glycosylated β-subunit. [14] In vascular smooth muscle, the occurrence of α1, α2 and α3 subunits has been reported in rat mesenteric artery, [9] rat aorta myocytes, [15] rat thoracic, superior mesenteric and tail arteries. [16] However, the role of sodium pump has not been characterized in the ruminal artery of the ruminant species. Similarly, there is limited information with respect to regulation of the ruminal arterial sodium pump by agonist. Using ruminal artery as a model for ruminant vascular smooth muscles, we employed isometric tension recording to study KCl-induced relaxation of goat ruminal artery rings contracted with K + -free solution and low concentration of 5-HT to study the functional regulation of sodium pump . There are several findings of the present study. First, endothelium of ruminal artery had no significant influence on the activation of sodium pump by extracellular K + and small increases in the extracellular concentration of K + (of < 30mM) caused concentration related relaxation of the goat ruminal artery in K + free medium, confirming that K + powerfully relaxes this artery. Second, this response to K + was significantly inhibited by 30 mM Ba 2+ , which, at this concentration, is considered a selective inhibitor of the inward rectifier K + (K ir ) channel.[17],[18] By contrast, ouabain had a significant inhibitory effect on K + -induced vasorelaxation in a concentration dependent manner, suggesting that the Na + -K + -ATPase has a potential role in ruminal vasodilatation. Third, digoxin at sub micromlar concentration (0.1µM) significantly inhibited the K + - induced relaxation at only 3 and 10mM. These observations clearly demonstrated the involvement of K ir channel and Na + -K + -ATPase in K + induced vasorelaxation of the goat ruminal artery. KCl-induced relaxation in vascular smooth muscles may involve several independent mechanisms, such as activation of sarcolemmal Na + -K + -ATPase and/or activation of inward rectifier K + -channels. [9] K + -induced dilation of small renal artery was attributed to the activation of smooth muscle Na + -K + -ATPase with no role for inward rectifier K + -channel. [19] One of the distinguishing features of vascular relaxation by K + involving Na + - K + -ATPase is that the extracellular concentration of K + is less than 5mM, whereas inward rectifier K + -channels primarily mediate K + -induced relaxation above the physiological K + -concentration (>5mM). [18] In goat ruminal arteries, we found that extracellular K + between 10µM and 10mM produced graded relaxation of the vessels bathed in K + -free solution and primed with 5-HT (0.1µM) to sustain the contractions. Activation of the K ir channel and conduction of an outward K + current in response to small increases in extracellular K + is thought to occur because of unique gating properties of K ir channels. [17],[18] Larger increases in K + (by < 30mM) cause smooth muscle depolarization and subsequent constriction of several arteries due to marked membrane depolarization and Ca 2+ entry via voltage-operated Ca 2+ channels. [18] Recent findings suggest that the K ir channel is indeed involved in mediating vascular smooth muscle hyperpolarization and vasorelaxation in response to K + . [19],[20] The present study provided evidence that K + - induced vasorelaxation in ruminal artery is Ba 2+ sensitive, and is likely to be at least partly mediated by activation of K ir channels. Ba 2+ (30mM) can selectively abolish smooth muscle relaxation in response to K + , whereas selective inhibitors of other K + channels do not inhibit K + -induced relaxation, [20] suggesting that these concentrations of Ba 2+ are sufficient to completely inhibit K ir channels and that K + is likely to activate only this type of K + channel. Interestingly, we observed only partial (31.7 ± 2.7%) inhibition of K + - induced vasorelaxation by Ba 2+ in this study, which is in contrast to findings of the studies in which Ba 2+ was reported to abolish completely the K + -induced vasorelaxation in blood vessels. [20] The discrepancy in the sensitivity to Ba 2+ of the K + -induced vasorelaxation versus hyperpolarization could be related to either altered vascular response specific to ruminal artery or an influence of several mediators that participate in ruminal contraction. When the ruminal arteries were immersed in a K + -free medium a small and transient contraction appeared. In other vessels, it has been reported that this response ranges from no increase in tension to a marked contraction [21],[22] suggesting different Na + pump activities in the vascular beds. These contractions are due to Na + pump inhibition produced by K + -omission. [23],[24] The subsequent addition of K + elicited vasodilatation which was blocked by ouabain more potently than digoxin. The inhibitory effect on K + -induced vasorelaxation was further blocked in presence of higher concentration (0.1µM) of ouabain. This finding clearly demonstrates that ouabain inhibits Na + pump in the ruminal artery in a concentration-related manner. Consistent with the expression of the sodium pump isoforms in rodents, recent studies on gene targeted mice emphasize a significant role of the a2 isoform in regulating contractility of blood vessels in vitro and regulation of blood pressure in vivo. [25],[26] The a1 isoform has been found to be having a "housekeeping role" in mouse aorta [26] and a2 isoform has been shown to possess a high affinity to low (submicromolar) concentrations of ouabain [27] in pulmonary vasculature. Thus far there is no information on the physiological roles of a1 and a2 isoforms in goat ruminal artery; the present functional study suggests that this isoform may have a role in regulating contractility of the ruminal vasculature In conclusion, the results of this study demonstrate that K + elicits marked relaxation of the goat ruminal artery. This effect may be (i) in part mediated through Ba 2+ sensitive Kir channel, as well as (ii) ouabain sensitive Na,K-ATPase isoform which may contribute to the vasorelaxant response. Further study is needed to confirm the molecular identity of Na,K-ATPase in goat ruminal artery. Acknowledgment The authors are thankful to Dr. S. K. Mishra, Ph.D, Principal Scientist, Division of Veterinary Pharmacology, IVRI, India for supplying gift sample of ouabain. References
Copyright 2009 - Indian Journal of Pharmacology The following images related to this document are available:Photo images[ph09020t1.jpg] [ph09020f1.jpg] |
| |||||||||

{kind=link}
{kind=link}