 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Tropical Journal of Pharmaceutical Research, Vol. 7, No. 2, June 2008, pp. 953-959 Research Article Uptake and kinetic properties of choline and ethanolamine in Plasmodium falciparum H Ahiboh1,3 , AJ Djaman2, FH Yapi2, A Edjeme-Aké1, ML Hauhouot-Attoungbré1, ED Yayo1, D Monnet1 1 Département de Biochimie Biologie Moléculaire, UFR des Sciences Pharmaceutiques et Biologiques, Université de Cocody – Abidjan BP V34 Code Number: pr08013 Abstract
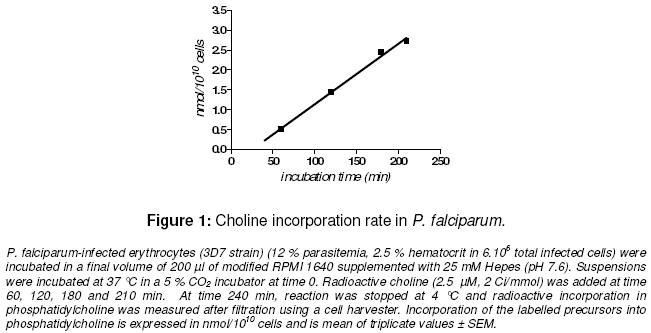
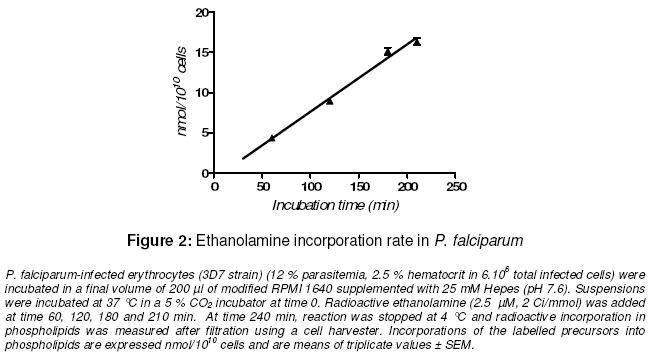
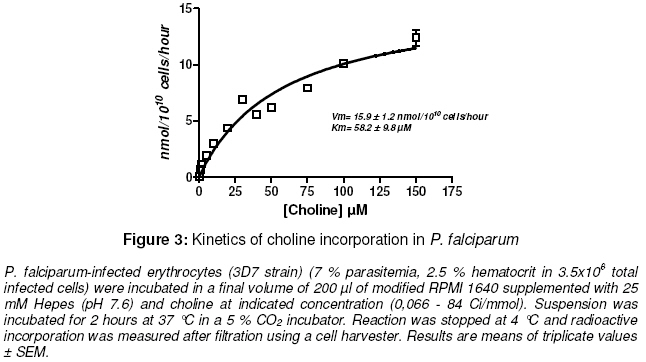
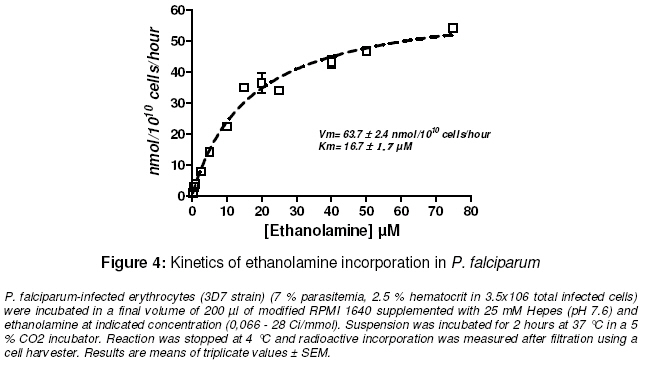
Purpose: The asexual proliferation of Plasmodium, inside the erythrocyte, is accompanied by the synthesis of huge quantities of phosphatidylcholine (PtdCho) and phosphatidylethanolamine (PtdEtn). These needful phospholipids for the cytoplasmic membrane of the merozoites are provided by the precursors choline and ethanolamine. PtdCho and PtdEtn are synthesized by the parasite because the erythrocyte is unable to do it. In order to assess the dynamism of the phospholipid pathways, we aimed to investigate the respective shape of the uptake of choline and ethanolamine by Plasmodium falciparum. Keywords: Plasmodium, phospholipids, kinetics, metabolism, choline, ethanolamine INTRODUCTION Like many other animal cells, the cytoplasmic membrane of the Plasmodium is made of bulk phospholipid molecules organized in a double layer. The major ones are phosphatidylcholine (PtdCho) and phosphatidylethanolamine (PtdEtn). After the invasion of the erythrocyte, the parasite starts a schizogonic cycle marked by the production of several merozoites. This asexual proliferation of the parasite inside the erythrocyte is accompanied by the synthesis of huge quantities of needful phospholipids for the membrane of the merozoites. It means a biosynthesis of PtdCho and PtdEtn by the parasite because the erythrocyte is unable to do it on its own. In order to assess the needs of phospholipids for the parasite, we evaluated the uptake of its precursors, choline and ethanolamine. Many studies reveal metabolic differences within the species of Plasmodium1-3 . The uptake of choline and ethanolamine by the Plasmodium seem to vary with the species, the stage of the life cycle and the experimental conditions4 . In this study, we determined the uptake and apparent kinetic properties of choline and ethanolamine in P. falciparum infestederythrocyte, the most widespread human malarial parasite in West Africa. EXPERIMENTAL In vitro culture of P. falciparum The 3D7 strain of P. falciparum in infected erythrocytes was incubated in modified RPMI 1640 medium supplemented with 25 mM Hepes (pH 7.6). The RPMI 1640 was free of choline and ethanolamine. The parasitemia was 7 -12 % with a hematocrit of 2.5 % in a total amount of 3.5 x106 – 6 x106 infected cells. The cell suspension was distributed in microwell plate and incubated at 37°C in a 5 % carbon dioxide atmosphere at time 0. Labelling for time-course studies To assess the uptake of the phospholipid precursors by the infected-erythrocyte, radioactive isotopes of [1-3 H]ethan-1-ol-2 amine and [methyl-3H]choline, purchased with Amersham™ corp were used They were added, in the respective microwells, at time 60, 120, 180 and 210 min (final concentration of 2.5 µM; specific activity of 2 Ci/mmol). The final volume of the suspension in each microwell was 200 µl. At time 240 min, reactions were stopped at 4°C. Radioactive incorporation in bulk molecules was measured after haemolysis, filtration and transfer onto glass-fiber filter plate using a cell harvester. Radioactivity was measured on a scintigraph counter from Beckman™. Incorporation of the labelled precursors into the infected erythrocyte was expressed in nmol/1010 cells. Labelling for kinetic assay To assess the kinetic features, different specific activity of radioactive choline and ethanolamine were added at indicated concentrations in the suspension of the infected-erythrocytes. The suspension was incubated for 2 hours at 37 °C in a 5 % CO2 incubator. Reactions were stopped at 4 °C and radioactive incorporation was measured after filtration and transfer on fiber glass plate using a cell harvester. Radioactivity was measured as described above. The blank test was performed with an uninfected-erythrocyte suspension provided by an uninfected human to assess nonspecific uptake by the erythrocyte. Data analysis Experiments were performed twice. Blank test was deduced from each result and all of them were means of triplicate assays. Regression curve was drawn and the Michaelis constant (Km ) and Vmax were calculated with the software GraphPad Prism® version 4. RESULTS Time course of choline and ethanolamine uptake P. falciparum incorporates choline and ethanolamine in its bulk molecules retained by the cell harvester. For the two precursors, the uptake is constant and linear at least from 60 to 240 minutes. The rate of choline uptake is 0.91 ± 0.03 nmol.1010 parasites.h-1 and 5.01 ± 0.24 nmol.10-10 parasites.h-1 for ethanolamine uptake (Figs. 1 and.2). The in vitro conditions show that the uptake of ethanolamine is greater than that of choline. Kinetics of uptake This assay was performed assuming an enzymatic process for the uptake of precursor and their incorporation in bulk molecules of the parasite. The uptake of choline and ethanolamine in P. falciparum showed an apparent Michaelis-Menten kinetics with the following features: choline uptake had a Vmax of 15.9 ± 1.2 nmol.10-10 cells.h-1 with an apparent K m of 58.2 ± 9.8 μM and ethanolamine uptake had a Vmax of 63.7 ± 2.4 nmol.10-10 cells.h-1 with an apparent Km of 16.7 ± 1.7 μM (Figs 3 and 4). The metabolic pathway of precursor incorporation in the phospholipids of P. falciparum seems to have a greater affinity for ethanolamine than for choline. Beside this, the incorporation of ethanolamine was faster than that of choline. DISCUSSION In human infection, signs of clinical morbidity, associated with malaria may occur at a parasitaemia range of about 100 – 720,000 per µl (i.e., 0.02 to 15%)5, 6. Our experimental conditions were chosen to get an optimal growth of the parasite. It means not too low parasitaemia to get a sufficient measurable metabolic reaction through the radioactivity signal. But not too high parasitaemia to get a dynamic and optimal growth of the parasite and avoid a high amount of lactic acid or other biological wastes that a high parasitaemia can release. It is the same caution that led to a relatively short incubation time (less than 240 min). In human blood, the choline concentration is about 10 -12 µM and ethanolamine is about 2.57 µM. In order to assess the match of choline and the ethanolamine pathways in in vitro conditions, we chose an identical final concentration of precursors (2.5 µM for each). The uninfected-erythrocyte is not expected to incorporate choline or ethanolamine in any metabolic pathways. However, Staines and Kirk showed that in an infected mammalian, the choline enters in uninfected erythrocyte with the same rate than in infected erythrocyte4 . Therefore we performed a blank test with uninfected-erythrocytes from healthy humans to deduce the non-specific adsorption of the precursors on any bulk molecules of the erythrocytes. Several pathways from choline or ethanolamine are known to lead to phosphatidyl choline and phosphatidyl ethanolamine. The one known as the Kennedy pathway, consists in the phosphorylation of choline in phosphocholine (PC), catalyzed by a choline kinase (EC 2.7.1.32). Then a cytidylmonophosphate (CMP) is transferred on the PC leading to CDP-Choline. This reaction is catalyzed by a cholinephosphate cytidyltransferase (EC 2.7.7.15). And finally, the CDP-Choline is transferred on a diacylglycerol (DAG) to get the PtdCho. This reaction is catalyzed by CDP-Choline:1,2 diacylglycerol choline phosphotransferase (EC 2.7.8.2). Another pathway goes through the tri-methylation of the phosphoethanolamine (PE) into the PC and then leads to PtdCho via CDP-Cho 8 . The last one, known as occurring in mammalian cells and yeast, is the tri-methylation of PtdEtn into PtdCho9 . The involvement of each pathway depends on the type of cell. The respective biosynthetic and catabolic pathways of phosphatidylcholine and phosphatidylethanolamine involve many kinds of molecules : choline is known to be mainly taken up and incorporated in phosphatidylcholine (PtdCho) without betaine or acetylcholine biosynthesis10 . Ethanolamine is retrieved from PtdCho and Phosphatidylethanolamine (PtdEtn). But nothing is known about the kinetics of their catabolism. Our experiments show that P. falciparum incorporates choline in a constant way within 60 – 240 minutes while P. knowlesi optimal incorporation of choline into PtdCho is less than 180 min. But the rate of choline uptake, supposed to be mainly incorporated in PtdCho, is about 100 fold lesser for P. falciparum than for P. knowlesi11 . In the same way, Staines and Kirk showed that the rate of choline uptake by P. vinckei is greater than that of P. falciparum one 4 . This correlates with the length of life cycle of P. knowlesi or P. vinckei which is 24 hours while P. falciparum is much longer, 48 hours. Infestation of the erythrocyte by Plasmodium entails a huge change of phospholipid composition in the erythrocyte membrane and in the whole complex parasite-erythrocyte relationship. At the schizonte stage phospholipid composition increases to about 500 – 700% and is mainly composed of PtdCho and PtdEtn (75 – 85%) with predominantly PtdCho12-14 . However in our condition of experiments, we noticed that the rate of ethanolamine uptake was superior to the rate of choline uptake. The normal mammalian plasma choline concentration is in the range 10 – 40 µM15 . Therefore, we expected the rate of choline uptake to increase and be superior to that of ethanolamine. But on the one hand Vmax of choline uptake was inferior to Vmaxof ethanolamine uptake. On the other hand, the apparent Km of ethanolamine uptake showed a higher affinity of the parasite for ethanolamine rather than choline. These observations contribute to the hypothesis that PtdCho is mainly issued from ethanolamine according the three steps of N-methylation of phosphoethanolamine to phosphocholine and catalyzed by phosphoethanolamine Nmethyltransferase (EC 2.1.1.103)8, 16 . Otherwise, in in vivo conditions, the ethanolamine pathway, is inhibited by a transcriptional repression of the phosphoethanolamine methyltransferase as recently shown by Witola and Ben Mamoun17 . This apparent feature of Mickaelis-Menten kinetics for the choline and ethanolamine uptakes hide the possibility that these pathways possess any key-steps regulated by a possible allosteric enzyme. CONCLUSION Choline and ethanolamine are incorporated in bulk molecules of P. falciparum. The uptake of these precursors by infected-erythrocyte increased when compared with uninfected erythrocyte. Such differences in the apparent kinetic properties of the choline and ethanolamine uptake are easily understandable if we are aware that ethanolamine has to provide both PtdCho and PtdEtn metabolic pathways. On the contrary, choline provides only the PtdCho pathway. These results are in favor for a most dynamic influx of ethanolamine in the P. falciparum phospholipids but may not be the case in in vivo conditions. ACKNOWLEDGEMENT The authors acknowledge Wein S, Maynadier M, Boudou F for their technical assistance. We thank Herbute S, Wengelnick K and Vial H at the Laboratory of “Dynamique des Interactions Membranaires Normales et Pathologiques” (CNRS UMR 5235) Université Montpellier II for their technical assistance and helpful advice concerning the protocol of experiments. This work was funded via an SCAC grant from the Cooperation Department of the French Ministry of Foreign Affairs. REFERENCES
© Pharmacotherapy Group, Faculty of Pharmacy, University of Benin, Benin City, Nigeria. The following images related to this document are available:Photo images[pr08013f3.jpg] [pr08013f2.jpg] [pr08013f4.jpg] [pr08013f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}