 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Tropical Journal of Pharmaceutical Research, Vol. 7, No. 4, December, 2008, pp. 1107-1116 Research Article Mathematical Modelling of Intraretinal Oxygen Partial Pressure R Avtar* and D Tandon Department of Mathematics, Harcourt Butler Technological Institute, Kanpur 208002, India Received: 15 July 2008 Revised accepted: 15 September 2008 Code Number: pr08032 Abstract Purpose: The aim of our present work is to develop a simple steady state model for intraretinal oxygen partial pressure distribution and to investigate the effect of various model parameters on the partial pressure distribution under adapted conditions of light and darkness.. Keywords: Mathematical modeling, Intraretinal oxygen pressure, Retinal capillaries, Oxygen consumption, Retinal vascular occlusion, Oxygen metabolism. INTRODUCTION Oxidative metabolism in the retina is a very important process and is an essential metabolic requirement of living and reproducing retinal cells as well as a necessity for the maintenance of ionic pump mechanisms in the retina which, in turn, maintains the integrity of retina . A proper oxygen environment (sufficient oxygen availability) is required for retinal oxidative metabolism. A physiological oxygen environment is maintained through a delicate balance of oxygen supply to the retina and oxygen consumption in different retinal layers of the retina. The rate of oxygen consumption is dependent on the rate of oxygen metabolism. The retinal layers, in which there is high rate of oxygen metabolism , have high rate of oxygen consumption . The oxygen metabolism and oxygen consumption are unevenly distributed in the retina. Any interference in this activity will cause tissue changes6 . The oxygen required for essential metabolism in the retina is primarily derived from the blood in choroidal vessels and in the central retinal artery. The choroidal blood vessels supply oxygen outer retina by diffusion retinal artery nourishes the oxygen enters the to the avascular whereas the central the inner retina. As retina, it creates a minimum tension/ partial pressure in the retina. This pressure is regulated through the balance of oxygen delivery and oxygen consumption in the retina2 . Besides, retinal blood flow is strongly dependent on the partial pressure of oxygen. Most of the oxygen delivered by choroidal circulation to the outer retina is consumed by photoreceptor segments because this layer is the location of all photoreceptor mitrochoridria with the exception of those in the synapse3 . A greater proportion of the oxygen provided by the retinal circulation to the inner retina is utilized by its inner plexiform layer. The rate of oxygen consumption varies from layer to layer and proportion of oxygen consumed by different retinal layers is not known. Over supply of oxygen can also result in some retinal diseases such as retinopathy. There is also evidence that oxygen toxicity plays a role in degenerative retinal diseases such as retinitis pigmentosa (RP)1 and it has been suggested that manipulation of the oxygen environment could be a therapeutic strategy in the management of retinal diseases such as RP, retinal detachment, and occlusive diseases of the retinal circulation. Even in healthy eyes, there is a very delicate balance between oxygen supply and oxygen consumption. Disruption of this balance may cause too little or too much oxygen to be present in specific retinal layers6 . Understanding the oxygen requirements of different components of retina is vital if therapeutic strategies to restore an appropriate oxygen environment are to be developed. Normal retinal oxygen environment (i.e., normal oxygen partial pressure in the retina ) is influenced by several physiological factors under normal and pathological conditions2 . Under a pathological state of central retinal occlusion which makes the whole retina anoxic, the blood supply to the inner retina is interrupted in proportion to the degree of occlusion and hence oxygen-delivery to the inner retina is affected. Under the normal characteristics of the choroid-retinal wall, the choroidal partial oxygen pressure, which ensures sufficient oxygen supply to the outer retina, has a dominating effect on the oxygen-environment. The states of normoxia, hyperoxia, extreme hyperoxia etc, affect retinal oxygenation. Light -adaptation and dark -adaptation of the retina also influence the oxygen partial pressure4 . The oxygen consumption in different retinal layers is bound to affect the pressure. The effect of such factors varies in different layers. The understanding of the effects of such factors on the oxygen partial pressure may be important in the understanding of retinal development and retinal pathology and in the clinical management of retinal vascular diseases. In order to devise strategies to restore optimal oxygen environment in the retina, there is a need to improve present understanding of retinal oxygenation and physiological factors affecting it. Enriched understanding may be useful in the development of more viable strategies for improving oxygen supply to ischemic retina. A mathematical model that accurately describes the intraretinal oxygen partial pressure distribution may contribute to improved understanding of retinal oxygenation. In 1999, Cringle et al.2 developed a steady state mathematical model for the oxygen partial pressure distribution in the avascular (outer) retina . The model treats the outer retina as consisting of four layers. The oxygen consumption in the outer photoreceptor layer and in the outer nuclear layer was considered to be zero, while in the inner photoreceptor layer and deep retinal capillaries, the oxygen consumption was considered to be constant. The effect of oxygen partial pressure levels under adapted light and dark conditions on the oxygen partial pressure in the retina was investigated and discussed. Linsenmeier and Silver3 presented a threelayered steady state model for the oxygen partial pressure distribution in the outer retina by assuming oxygen diffusion and constant oxygen consumption inside it. They fitted their experimental data to the model. In 2002, Cringle et al1 generalized their earlier model and presented an eightlayered steady state model in the retina. The oxygen consumption in different layers was considered to be constant. They computed oxygen tension in different layers of the retina and compared the model’s results with experimental results. Roos4 presented a transient four-region model for the oxygen partial pressure in the retina. The model divides the outer retina into three regions and the inner retina was considered as the fourth region. The oxygen-consumption was assumed to follow Michaelis-Menten kinetics and oxygen delivery in the inner retina was described using Hill’s equation and Fick’s law. They investigated the effect of changes in retinal blood flow under the light and dark adapted conditions. The objective of the present study is to formulate a simple steady-state mathematical model for intraretinal oxygen partial pressure distribution. The model treats the retina as consisting of eight layers. The oxygen consumption was assumed to follow Michaelis-Menten kinetics and oxygen delivery in the inner retina was described by using Hill’s equation and Fick’s law4 .The approximate solution to the model was obtained using an iterative technique and Runge-Kutta Nystrom’s method5 . The computational results of the model have been presented in the Figures and are also discussed. METHODS Mathematical model We considered a retina that has eight layers (see Fig 1). Layers 1-3 constitute the outer retina and the remaining layers constitute the inner retina. The outer retina is avascular and hence does not receive oxygen. Layer 2 is the location of a majority of the photoreceptor mitrochondria. Most of the oxygen consumption in the outer part of the retina seems to take place in the photoreceptors’ inner segments1 . In the inner retina (which is vascular), oxygen transport to the retinal cells occurs via blood and by diffusion. Most of the oxygen supply in the inner part of retina seems to take place in the layer 61 . The oxygen consumption and oxygen delivery was assumed to occur in the outer region of the inner plexiform layer1 . Boundary and interface conditions The physiologically relevant and mathematically consistent boundary and interface conditions are prescribed below: In the above equations, x represents the distance from the choriocapillaries. Eq.5 represents that the partial pressure of oxygen at the choridal retinal boundary and is equal to that in the choriocapillaries. Eqs 6(a,b), 7(a,b), 8(a,b), 9(a,b), 10(a,b), 11(a,b) and 12(a,b) represent the continuity of both partial pressure of oxygen and flux at the junctions between adjacent layers of the retina. Eq (13) prescribes absence of oxygen flux at the retina – vitreous boundary. Assumptions To this end, the following assumptions are introduced:
Governing equation The steady state local partial pressure of oxygen in the retina is governed by the diffusion equation:
In view of the above-mentioned assumptions, eq. (1) reduces to a one dimensional form:
where Pi is the partial pressure of oxygen in the ith layer, Dithe diffusion coefficient of oxygen in the ith layer, qi the oxygen consumption term in the ith layer, si the oxygen delivery term in the ith layer. According to Michaelis-Menten kinetics, the consumption term for oxygen [16] is given by:
where (qox max )i is the maximal rate of oxygen consumption in the ith layer and ki the partial pressure of oxygen at half maximal
consumption speed in the ith layer. Since a greater proportion of the oxygen consumption occurs in the layer 2, therefore, maximal oxygen consumption rate in this layer will be 90mmHg−1 in light and 170mmHg−1 in dark. For the inner retina, the value of maximal oxygen consumption rate will be 26 mm Hg −1 in both light and dark which is a value within the range reported by others5,6 . In layers 1, 3, 5, 7 and 8, oxygen consumption was assumed to be zero. The oxygen consumption in the outer retina was confined to the inner segment of the photoreceptor layer. In the inner retina, oxygen consumption was assumed to occur in the outer region of the outer plexiform layer. The amount of oxygen transported locally from the blood to tissue is given by4 :
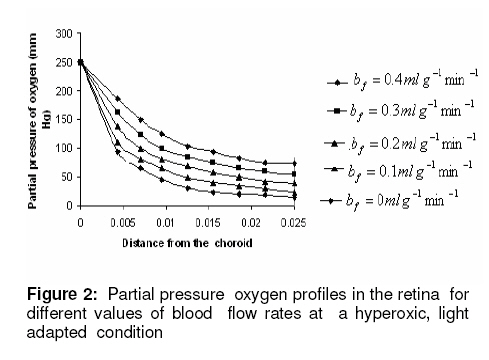
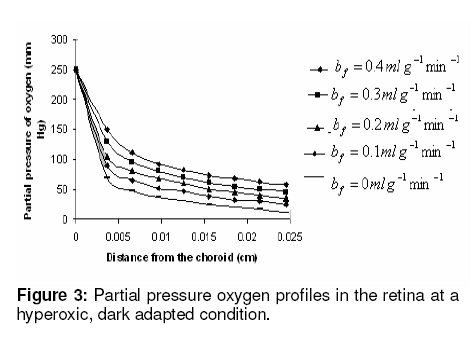
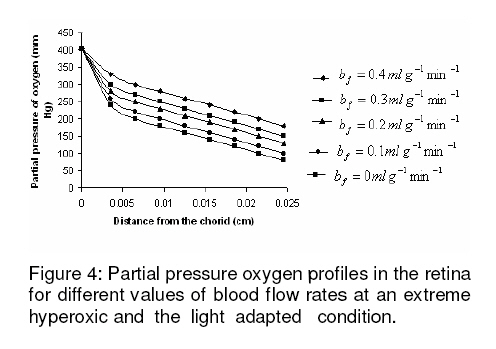
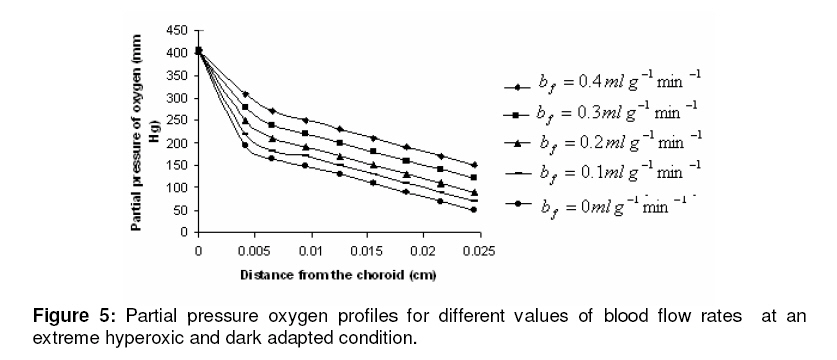
where β is a constant , pi the local partial pressure of oxygen in the ith layer , pblood the partial pressure in arterial blood, Hb the hemoglobin concentration in blood, α1 the solubility of oxygen in blood, δ the oxygen carrying capacity of hemoglobin, n Hill coefficient and bf blood flow rate in the inner retina. In the outer retina oxygen delivery was assumed to be zero since the outer retina does not have any blood flow. In the inner retina, oxygen delivery occurs only in the outer region of the outer plexiform layer . Solution to the model The system of second-order differential equations governing the partial pressure of oxygen in different layers of the retina is nonlinear, due to the presence of non-linear consumption term in the equation for po 2 in the second layer and non-linear consumption and delivery terms in the equation for po 2 in the sixth layer. The system of equations is linearized by considering zero order consumption of oxygen in the second and the sixth layers and non-delivery of oxygen to the sixth layer, and so approximate solution to the system of equations is obtained. In order to improve the approximate solution for po 2 in the second layer, we determine p2 and RESULTS The computational results of the model for the partial pressure of oxygen in the retina were obtained by using typical values of model parameters for a representative eye given in Table 1. The distributions of oxygen partial pressure in a hyperoxic and light adapted condition for different values of blood flow rate in the inner retina are shown in Fig 2. It is evident from the plots in Fig 3 that the partial pressure of oxygen decreases along the retinal depth. The partial pressure of oxygen is maximum at the choroidal side and decreases sharply in the outer and inner photoreceptor layers. This occurs as a result of the oxygen consumption in the inner photoreceptor layer. Due to the oxygen consumption and oxygen supply in the inner plexiform layer, there was less reduction in the remaining layers. It was observed from the plots that as blood flow rate in the inner retina increased, the partial pressure of oxygen increases. The oxygen partial pressure profiles in the retina at a hyperoxic and dark adapted condition are shown in Fig 3. The various plots in Fig 4 show that the partial pressure of oxygen decreases along the retinal depth. It is obvious from the plots in Figs 2 and 3 that the partial pressure of oxygen is higher in light than in the dark. The various oxygen partial pressure profiles in extreme hyperoxic, light adapted condition for different values of blood flow rate in the inner retina are depicted in Fig 4. From the plots in Fig 4, it can be appreciated that the partial pressure of oxygen decreases gradually across the retinal layers from the choroid to the vitreous body. A higher decrease in the partial pressure of oxygen in the inner photoreceptor layer and its adjoining layers than the outer photoreceptor layer and outer nuclear layer was observed. This decrease in oxygen partial pressure reduced the partial pressure of oxygen in the inner retina. The oxygen partial pressure profiles at an extreme hyperoxic and dark adapted condition are depicted in Fig 5. The different plots in Fig 5 demonstrate that under extreme hyperoxia condition the partial pressure of oxygen is extremely high at the choroidal side and decreases sharply along the retinal depth. The oxygen consumption and oxygen delivery occurring in the inner retina diminish the decrease in the retinal layers beyond inner plexiform layer as is evident from the plots in Fig 5. Comparison of the plots in Fig 4 and 5 show that the partial pressure of oxygen at an extreme hyperoxic and light adapted condition is higher than that in extreme hyperoxic and the dark adapted condition. DISCUSSION An insufficient supply of oxygen to the retina is thought to be an important pathogenic factor in a variety of retinal diseases such as retinal pigmentosa, retinal artery occlusion, etc1 . Oversupply of oxygen can also result in retinal disease4 . There is also evidence that oxygen toxicity plays a role in degenerative retinal diseases and it has been suggested that manipulation of the retinal oxygen environment may be a therapeutic tool in the management of retinal diseases 1,6 . Previous theoretical modeling of retinal oxygenation has mostly been focused on conditions where oxygen availability does not limit the oxygen consumption 4,5 . Moreover, attention has been focused on the outer retina or conditions where there is no retinal blood supply. However, Cringle and Yu1 have sought to include the inner retina with a possible blood circulation by introducing their oxygen consumption model. In their model, the layers were assumed to have blood supply, and the oxygen delivery and consumption were treated as a lumped term for oxygen supply and consumption, and a curve-fitting routine was used to obtain the parameter values producing the best fit to experimental data. This means that their model should be used for the interpolation of some aspects of experimental data. Thus, the present work has provided theoretically derived values of po2 in the retina at different degrees of retinal ischemia in light and darkness. If the result of this work is accepted as reasonable, it seems that more clinical studies with different degrees of hyperoxia will be required. CONCLUSION The computational results of the model predict that the partial pressure of oxygen in different layers of the retina is reduced by a decrease in the blood flow rate in the inner retina. The pressure is observed as minimum when there is no blood flow in the inner retina. This minimum pressure may fall below the critical level of oxygen partial pressure and affect the retinal function. In order to restore normal retinal function, extreme hyperoxia may assist the choroid to supply oxygen to the whole retina during total retinal artery occlusion as suggested by Roos4 . The results also predict higher partial pressure of oxygen in the retina under light adapted conditions than that under dark adapted conditions. Thus, light may, to some extent, be beneficial in preventing ischemia caused by retinal artery occlusion, etc. ACKNOWLEDGMENT The authors gratefully acknowledge the constructive and fruitful comments of the reviewers of original manuscript of the paper. REFERENCES
© Copyright 2008 - Pharmacotherapy Group, Faculty of Pharmacy, University of Benin, Benin City, 300001 Nigeria. The following images related to this document are available:Photo images[pr08032f4.jpg] [pr08032f3.jpg] [pr08032f2.jpg] [pr08032f1.jpg] [pr08032f5.jpg] [pr08032t1.jpg] |
| |||||||||

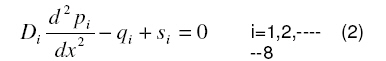
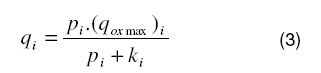
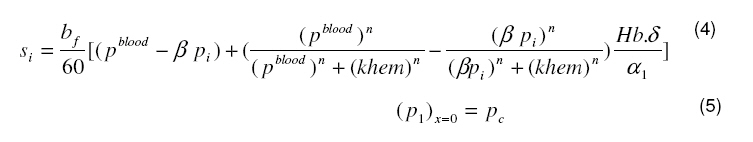
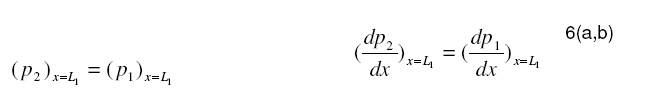
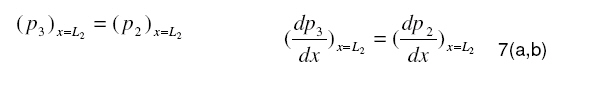
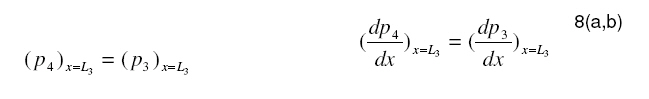
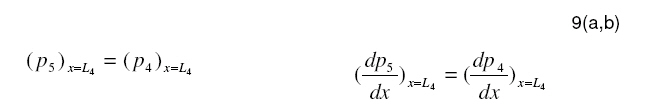
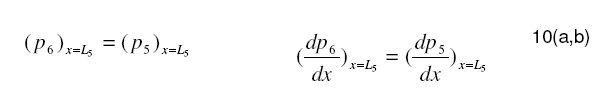
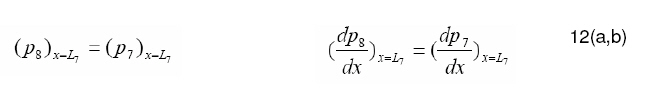

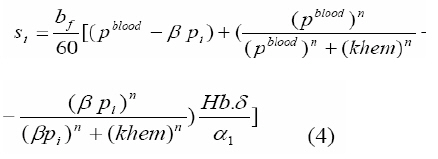
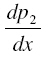 at x = L1from the solution for po 2 in the first layer through the interface condition 6(a,b) which serves as initial condition when we solve the nonlinear equation for po 2 in the second layer based on Runge-Kutta Nystrom’s method. In order to find an improved solution for po 2 in (i + 2)th layer, (i=1,2—6) we determine ( p) x=Li+1 and
at x = L1from the solution for po 2 in the first layer through the interface condition 6(a,b) which serves as initial condition when we solve the nonlinear equation for po 2 in the second layer based on Runge-Kutta Nystrom’s method. In order to find an improved solution for po 2 in (i + 2)th layer, (i=1,2—6) we determine ( p) x=Li+1 and 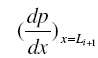 from the solution for po 2 in (i + 1)th layer as initial conditions when we solve differential equation for po 2 in (i + 2)th layer by Runge-Kutta Nystrom’s method. This procedure is repeated until
from the solution for po 2 in (i + 1)th layer as initial conditions when we solve differential equation for po 2 in (i + 2)th layer by Runge-Kutta Nystrom’s method. This procedure is repeated until 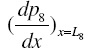 is less than a prescribed tolerance ( 10−5 ) or tends to zero. Here, three iterations are sufficient to yield satisfactory result for all the cases under consideration.
is less than a prescribed tolerance ( 10−5 ) or tends to zero. Here, three iterations are sufficient to yield satisfactory result for all the cases under consideration. 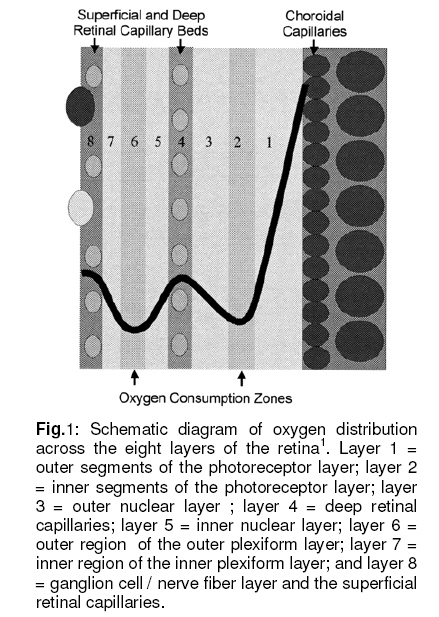{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}