 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Tropical Journal of Pharmaceutical Research, Vol. 8, No. 4, Aug, 2009, pp. 345-352 Research Article Antiproliferative Effect of the Methanol Extract of Piper crocatum Ruiz & Pav Leaves on Human Breast (T47D) Cells In-vitro Britanto Dani Wicaksono1, Yohana Ayupriyanti Handoko2, Enos Tangke Arung1,2, Irawan Wijaya Kusuma2, Dina Yulia1, Aldrin Neilwan Pancaputra3 and Ferry Sandra1* 1Stem Cell and Cancer Institute, Jalan Ahmad Yani no. 2Pulomas,
Jakarta, 13210, 2Wood Chemistry Laboratory, Forest Product
Department, Forestry Faculty, Mulawarman University; Jalan KH. Dewantara,
Kampus Gn. Kelua, Samarinda, East Borneo 75123, 3Dharmasis Cancer
Hospital, Jalan Letjend S. Parman Kav 84-86, Jakarta 11420, Indonesia. Received: 6 February 2009 Code Number: pr09045 Abstract Purpose: To
investigate the anti-cancer properties of the methanol extract of Piper
crocatum Ruiz & Pav. leaves and its mode of action in human breast
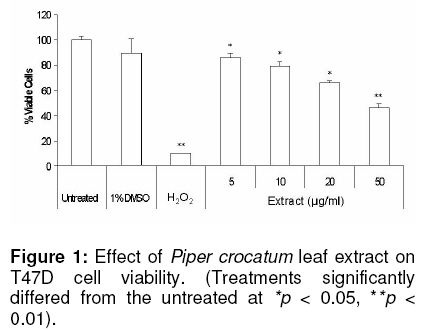
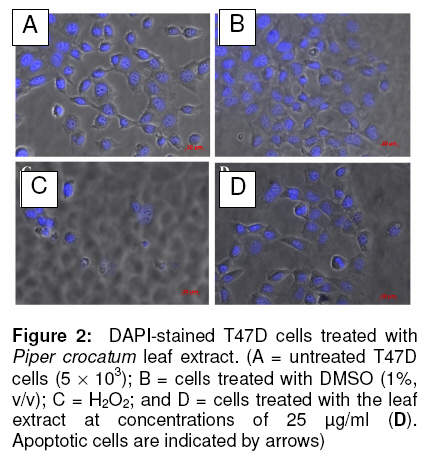
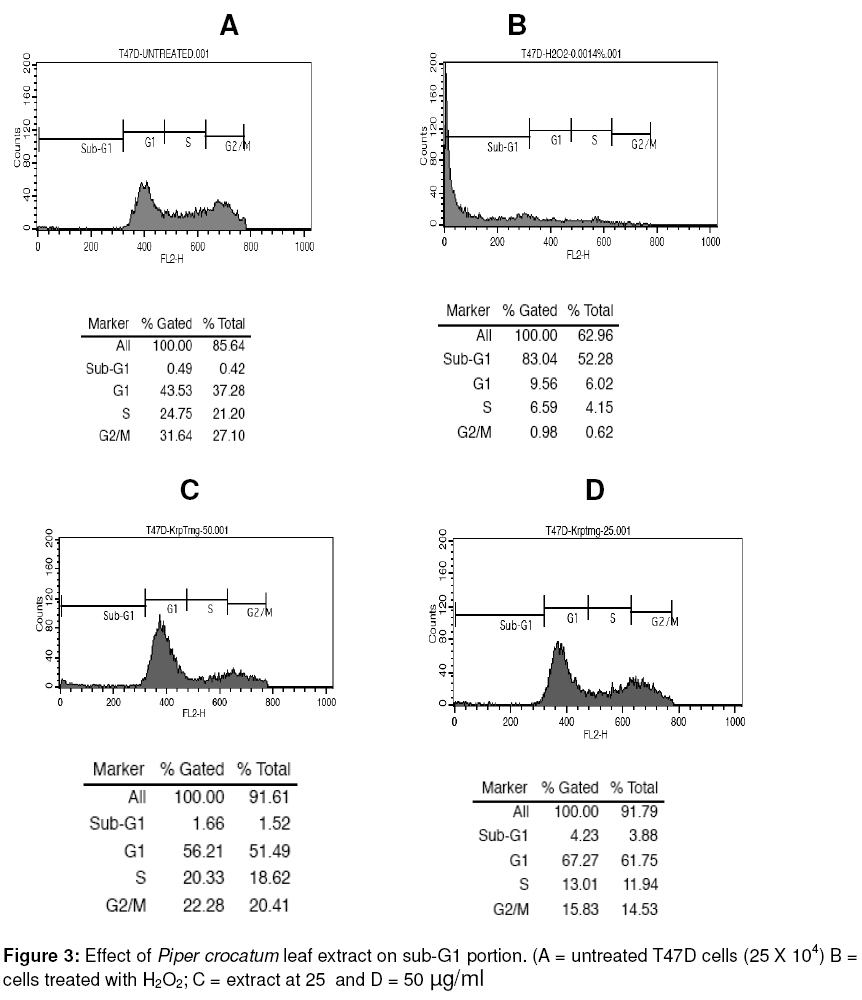
cancer (T47D) cells. Keywords: Piper crocatum leaf; methanol extract; antiproliferative; breast cancer; T47D cells INTRODUCTION Silymarin is an active extract of the seeds of Cancer is a major health problem and a leading cause of death not only in developed countries, but also in developing countries where approximately 72 % of all cancer death in the world occurred. Among women, breast cancer is the most commonly diagnosed cancer and frequent type of cancer causing death, resulting from metastatic spread of primary tumors1. Although common methods for cancer treatment have yielded some advantages, there is an ongoing need for both improvement of current therapeutic strategies and the search for novel agents. Throughout medical history, nature has long been shown an excellent and reliable source of new drugs, including anti-cancer agents. Approximately more than 60 % of currently used anticancer chemotherapeutics are derived in one way or another from natural sources, including plants2,3. In addition, there are more than 85,000 plant species that have been documented for medical use globally. The World Health Organization (WHO) estimates that almost 75 % of the world’s population has therapeutic experience with traditional remedies4. The major categories of plant-derived compounds that have medicinal properties are terpenoids, flavonoids, and alkaloids5. The origin of several cancer therapies currently being used are traceable to plants. Examples are vincristine and vinblastine from Catharanthus roseus6, taxol and docetaxel from Taxus brevifolia7, and campthotecins from Camptotheca acuminata8. Piper crocatum Ruiz and Pav. is traditionally used by Indonesians for treating various diseases, including breast cancer9. In East Borneo, from where the specimen for this study was collected, this plant is called Kerapakng Timang and people have empirically used it for curing cancer, which is traditionally named a deadly disease. To the best of our knowledge, however, there is no scientific evidence currently available regarding the anti-cancer properties of this plant. This, therefore, prompted us to investigate the anticancer properties of the methanol extract of the leaf of this plant on human breast cancer (T47D) cells as a model system. The biological activity of the extract was investigated in vitro by investigating cell viability, cellular and nuclear morphology, and examining sub-G1 portion after treatment with the extract. In addition, a study was also conducted to detect changes in the activity level of p44 and p42 mitogen-activated protein kinases (MAPKs; also known as extracellular signal-regulated protein kinases 1 and 2 (ERK1/2)) when cells were exposed to the extract. The p44/p42 signaling was investigated since this signaling pathway has been associated with cell growth and represents an important target for anti-cancer therapy10,11. Some plant-derived products have also been reported to alter p44/p42 activities12-14. EXPERIMENTAL Plant extract The leaves of Piper crocatum Ruiz & Pav. were collected from East Borneo forest by an exploration team from the Wood Chemistry Laboratory, Forestry Faculty, Mulawarman University, Samarinda, East Borneo, Indonesia and Stem Cell and Cancer Institute (SCI) Jakarta, Indonesia on March 2008. The plant was identified by Raharjo, of Dendrology Laboratory, Forestry Faculty, Mulawarman University, Indonesia and the voucher specimen (SCI-56) was deposited in the Wood Chemistry Laboratory, Forestry Faculty, Mulawarman University, Indonesia. The air-dried and ground leaves (10 g) were soaked in methanol (250 ml) for 24 h, and then the mixture was filtered to remove the marc. The extract was concentrated using rotary evaporator under vacuum at room temperature to obtain a final residue (1 g) for further experiment. The extraction was conducted at the Wood Chemistry Laboratory, Forest Product Department, Forestry Faculty, Mulawarman University, Samarinda, East Borneo, Indonesia. Cell culture T47D cells were obtained from the Indonesian Institute of Sciences Research Centre for Chemistry, Natural Products, Food and Pharmaceuticals Division, Bandung, Indonesia. The cells were grown and maintained in Dulbecco’s Modified Eagle’s Medium (DMEM) with L-glutamine supplemented with 10 %v/v fetal bovine serum, sodium bicarbonate, 100 µg/ml streptomycin and 100 U/ml penicillin at 37oC in humidified atmosphere of 5 % CO2. Cell viability assay Cell viability was determined by a microculture tetrazolium teachnique (MTT) assay performed according to the method described by Arung et al15, with minor modifications. The MTT assay provides a quantitative measurement of viable cells by determining the amount of formazan crystals produced by metabolically active cells. Briefly, cells were seeded into a 96-well plate (5 x 103 cells per well). After 24 h incubation, the medium was replaced with fresh medium containing leaf extract, dimethyl sulfoxide (DMSO; 1 %v/v in water), hydrogen peroxide (H2O2; 0.0014 %v/v in water) or medium alone and the cells were incubated for a further 24 h. DMSO (1 %v/v in water) was used as a negative control since this was used to re-suspend the extract. MTT reagent [3-(4,5-dimethyl-2.thiazolyl)-2,5-diphenyltetrazolium bromide (10 µl) in PBS (5 mg/ml)] was added to each well. The plate was incubated in a humidified atmosphere of 5% CO2 at 37oC for 3 h and then the medium was discarded and formazan crystals were dissolved in 1.0 ml of 0.04 N HCl and incubated for 2 h. The absorbance of cells was measured at 570 nm by a microplate reader. Untreated cells were counted with a hemacytometer and used for interpolating the absorbance of MTT assay results. The data was presented as percentage of viable cells (%). The data was analysed with the two-tailed student’s t-test against the untreated sample. Structural analysis and DAPI staining DAPI staining was performed as described by Sandra et al16 to determine the level of apoptosis in the cells. Briefly, the cells (5 ´ 103) were seeded onto sterilized cover slips in 60 mm Petri dish for 24 h and treated with the extract for another 24 h. Untreated cells and cells treated with DMSO (1%, v/v) and H2O2 (0.0014%, v/v) were used as controls. Treated cells and controls were rinsed with phosphate buffered saline (PBS), fixed with ice-cold 10% trichloroacetic acid, and further washed with cold 70, 80, 90 % and absolute ethanol. The cells were permeabilised with Triton-X (10 %v/v) and stained with 1 µg/ml 4’-6-diamidino-2-phenylindole (DAPI) for 4 min. To reduce the background, the stained cells were washed with PBS and were cover-slipped with 90% glycerol and observed under a fluorescence microscope (Zeiss Axio Observer Z1, Göttingen, Germany). Sub-G1 apoptosis assay and flow cytometry The cells were separated into four treatment groups as mentioned in the cell viability assay section above and analyzed for sub-G1 apoptosis using the method described by Sandra et al16. Briefly, the cells were seeded into a 24-well plate (25 x 103 cells per well) and incubated for 24 h. The medium was then replaced with fresh medium containing the extract, DMSO, H2O2 or medium alone. After 24 h, the cells were harvested and suspended in 1 ml of hypotonic fluorochrome solution (50 µg/ml propidium iodide in 0.1% sodium citrate containing 0.1% Triton X-100). The cell suspensions were placed in the dark for 1-2 h at 4oC before flow cytometric analysis. The propidium iodide fluorescence of individual nuclei was measured using FACS Calibur facility (Becton Dickinson, San Jose, California). Immunoblotting Western blot was performed as described previously by Sandra et al16. For immunoblotting, the various cell treatments investigated were: (i) cells treated with the extract (50 µg/ml) for 2 h; (ii) cells treated with insulin (1µg/ml) and harvested after 10 and 30 min later; and (iii) cells treated with the extract (50 µg/ml) for 2 h in addition to insulin (1µg/ml) observed at 10 and 30 min after insulin treatment. Untreated cells were also observed as negative control. The untreated and treated cells were lysed with buffer [20 mM HEPES buffer pH 7.2, 150 mM NaCl, 2 mM EDTA, 1 % Triton X-100, 50 mM sodium fluoride, 2mM sodium orthovanadate, 30 mM sodium pyrophosphate and a cocktail of protease inhibitors (containing 10 µM pepstatin A, 10 µM leupeptin and 1 mM p-amidinophenyl methanesulfonyl fluoride; Sigma)] and scraped. Protein concentration was determined using the Bio-Rad protein assay kit (Bio-Rad, CA, USA). Proteins from cells were separated by 10% sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a polyvinylidene difluoride (PVDF) membrane. Equal loading of proteins was routinely ascertained by Ponceau S staining. After blocking with 5% skim milk in Phospate-bufferred saline (PBS, pH 7.2), the membrane was incubated in 1 : 1,000 diluted rabbit polyclonal anti-phospho-p44/42 MAP Kinase (Thr202/Tyr204; Cell Signalling Technology, MA, USA), or 1 : 1,000 diluted mouse monoclonal anti-β-actin antibodies (Santa Cruz Biotechnology, CA, USA). The secondary antibody used was 1: 1,000 diluted horseradish-peroxidase-conjugated goat anti-rabbit (Cell Signalling Technology, MA, USA) or 1:2000 diluted anti-mouse (Santa Cruz Biotechnology, CA, USA) IgG antibodies. The bound antibodies were visualized by G:Box Syngene System (Beacon House, Cambridge, UK). Statistical analysis The IC50 (median inhibition concentration) is the concentration of toxic compound that reduces the biological activity by 50 %. The IC50 value was obtained from the MTT assay and calculated using non-linear regression analysis in Microsoft Excel software. The value was expressed as a geometric mean. Differences were considered to be statistically significant when p < 0.05 and p < 0.01. RESULTS Cytotoxic effects of leaf extract in T47D cells The results of the assessment of the cytotoxic effect of various extract concentrations of Piper crocatum leaf (5, 10, 20 and 50 µg/ml) on T47D cells after 24 exposures, determined by MTT assay, are shown in Figure 1. Cells with media only (untreated) and with added DMSO were also measured as negative controls, while cells treated with H2O2 was used as a positive control. A concentration-dependent decrease in cell viability was observed due to extract treatment, while only a small decrease (< 20%) in cell viability occurred when cells were treated with DMSO, the solvent used to re-suspend the extract. IC50 was reached at a concentration of 44.25 µg/ml. Cell and nuclear morphology Based on MTT assay results, DAPI staining was conducted to investigate whether the leaf extract reduced cell viability by inducing apoptosis, which is indicated by the appearance of nuclear fragmentation and changes in cell morphology. As seen in Figure 2, the cells with no extract treatment (Figure 2 A) and cells treated with DMSO and the extract (Figures 2 B and D) showed similar cell and nuclear morphology, and apparently, there was no indication of apoptosis. On the other hand, the cells treated with H2O2 (Figure 2 C) showed nuclear fragmentation and cell morphology changes, indicating apoptosis (indicated by arrows). Although there was no apoptosis, it appeared that the extract inhibited the growth of the cells (compare Figure 2 A and B with Figure 2 D). Sub-G1 apoptosis To further verify that the leaf extract did not cause cell apoptosis, flow cytometric analysis of cells stained with propidium iodide was conducted. The results confirmed that there was no nuclear fragmentation, indicative of apoptosis, in cells as shown by negligible changes in sub-G1 portion after treatment with the extract. Sub-G1 fraction in the cells without treatment was 0.49 % (Figure 3 A), while those treated with H2O2 (positive control) was 83.04 % (Figure 3 B). After treatment with 25 and 50 µg/ml of the extract, sub-G1 fractions formed were 1.66 and 4.23 %, respectively (Figures 3 C and D). Immunoblotting Since leaf extract apparently may not cause apoptosis, the mechanism of the antiproliferative effect of the extract was investigated by examining the expression of activated p44/p42 proteins. Transmission of insulin signal in cell proliferation was detected also via p44/p42 activation (Capeau, 2003), and therefore, treatment cells with insulin were also conducted as a control. Figure 4 shows that T47D cells exposed to 50 µg/ml of the leaf extract appeared to decrease p44/p42 activities compared to the untreated, as indicated by almost undetected signal intensity in the treated cells, while weak signals appeared in the untreated. Cells treated with insulin at 1 µg/ml caused transient p44/p42 activation, which confirms the involvement of p44/p42 in transmitting insulin signal (Figure 4). Inclusion of 50 µg/ml of the leaf extract in the incubation medium decreased expression of insulin-stimulated phosphorylation of p44/p42 at 10 and almost no signal was detected after 30 min (Figure 4). DISCUSSION Today, cancer remains one of the leading causes of death around the globe. Various cancer therapies have currently been tried, including the use of natural products from higher plants. Therefore, there is an urgent need to discover an effective, novel and scientifically reliable natural compound. Furthermore, an understanding of the mechanism of an anti-cancer agent is also important for future therapeutic application. The extract reduced cell viability in a concentration-dependent fashion as shown in the MTT studies. Proliferation of T47D cells exposed to the leaf extract was potently inhibited compared to the unexposed cells, based on microscopic studies (data not presented in this work) and no cell apoptosis was observed as examined by DAPI staining and sub-G1 apoptosis assay using flow cytometry. Intracellular target of methanolic extract of P. crocatum leaves was further explored by examining the activity level of protein tyrosine kinase, p44/p42. Denhard10 and Seger and Krebs11 explained that p44/p42 is a central player in Ras/MAPK cascade that regulate cell growth, differentiation and survival. T47D cells exposed to 50 µg/ml of P. crocatum leaf extract decreased p44/p42 activities. This result is similar to that reported by Daniels et al12 which indicated that aqueous and ethanol extracts of Anemopsis californica at a concentration of 50 µg/ml decreased ERK1/2 activities in human MCF-7/AZ breast and HCT8/E11 colon cancer cells. This activity might be responsible for the growth inhibitory effect of the extract in the cancer cells tested. Van Slambrouck et al14 also showed that cell growth was inhibited and levels of ERK1/2 were decreased when MCF-7/AZ breast cancer cells were exposed to the aqueous extract of J. communis. Hollosy and Keri18 also reported that many natural protein tyrosine kinase inhibitors have been derived from plants. The transmission of insulin signal in cell proliferation was known via p44/p42 activation17. This study shows that insulin transiently stimulated p44/p42 activation. This result supports the data previously reported by Pawelczyk et al19. They reported that exposure of lymphocytes to insulin produced a transient increase in ERK1/2 phosphorylation, indicated by a significant increase of activated ERK1/2 at 1 and 5 min, and then a decrease at 15 and 30 min. To investigate whether the P. crocatum leaf extract have consistent inhibitory effect on p44/p42 phosphorylation, we examined the response of insulin-stimulated phosphorylation of p44/p42 to extract treatment. The data showed that apparently, the extract inhibited activation of insulin-stimulated phosphorylation of p44/p42, indicated by decreasing phosphorylation of these proteins after 10 min and virtual absence of a detectable signal after 30 min. These data confirm that P. crocatum leaf extract inhibited proliferation of T47D cells via p44/p42 pathway. CONCLUSION The results suggested that Piper crocatum Ruiz & Pav. leaf extract inhibited proliferation of human breast cancer (T47D) cells. No apoptosis was observed due to Piper crocatum leaf extract treatment. Intracellular mechanism examination suggested that the extract inhibited cancer cell growth via p44/p42-dependent way. Components consisted of this extract, therefore, may have promise for use as a cancer chemotherapeutic agent in the future and can be used as scientific evidence to support anti-cancer properties of Piper crocatum. ACKNOWLEDGEMENT We acknowledge Ms Tjandrawati Mozef of the Indonesian Institute of Sciences Research Centre for Chemistry, Natural Products, Food and Pharmaceuticals Division, Bandung, Indonesia for generosly donating T47D cells. REFERENCES
© Pharmacotherapy Group, Faculty of Pharmacy, University of Benin, Benin City, 300001 Nigeria. The following images related to this document are available:Photo images[pr09045f3.jpg] [pr09045f4.jpg] [pr09045f2.jpg] [pr09045f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}