 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Tropical Journal of Pharmaceutical Research, Vol. 8, No. 4, Aug, 2009, pp. 353-360 Research Article Production of Extracellular Anti-leukaemic Enzyme L-asparaginase from Marine Actinomycetes by Solid-state and Submerged Fermentation: Purification and Characterisation N Saleem Basha1*, R Rekha2, M Komala1 and S Ruby3 1Department of Pharmaceutical Biotechnology, 2Department
of Pharmacognosy, 3Department of Pharmaceutical Chemistry, Mohamed Sathak A J College of Pharmacy, Medavakkam, Chennai 600 119, India. Received: 23 November Code Number: pr09046 Abstract Purpose: The objective of this investigation was to isolate marine
actinomycetes, screen them for L-asparaginase activity and characterise the
enzyme. Key words: L-asparaginase, Solid-state media, TGY broth, TFY broth, SDS-PAGE, Km and Vmax. INTRODUCTION Marine microbes represent a potential source for commercially important bioactive compounds1,2. Among marine microorganisms, actinomycetes have gained special importance as the most potent source of antibiotics and other bioactive secondary metabolites3. While most of the studies on actinomycetes have focused on antibiotic production, only few reports have dwelt on their enzymatic potential4. The discovery of L-asparaginase (L-asparaginase aminohydrolase, E.C.3.5.1.1), a medicinal agent for the treatment of malignant tumors, was made in 1922 5. Clementi showed that guinea pig serum contained a high activity of L-asparaginase5, while Mashburn and Wriston successfully purified Escherichia coli L-asparaginase and demonstrated its tumour-inhibitory activity5. L-asparaginase converts L-asparagine to L-aspartic acid. Since several types of tumour cells require L-asparagine for protein synthesis, they are deprived of an essential growth factor in the presence of L-asparaginase. Effective depletion of L-asparagine results in cytotoxicity for leukaemic cells5 but thus far, tumour inhibitory activity has been demonstrated only with asparaginases from E. coli, Erwinia aroideae and Serratia marcescens6. The administration of such an enzyme protein for a long duration, in general, produces the corresponding antibody in the tissues, resulting in anaphylactic shock or neutralization of drug effect. Therefore, the use of new serologically different L-asparaginase with a similar therapeutic effect is highly desirable7. Hitherto, L-asparaginase was produced by the technique of submerged fermentation. However, solid-state fermentation (SSF) is a more effective technique as the yield of the product is several times higher than that of submerged fermentation (SF). It also offers many other advantages including resistance to contamination, ease of product extraction and simpler methods for treating the fermented residue8. The objective of this study was to isolate L-asparaginase from marine actinomycetes by solid-state fermentation using soybean meal and submerged fermentations well as partial purification, and characterization of the crude enzyme extract EXPERIMENTAL Materials Marine sediment samples were collected from different sites in Tamilnadu and Kerala, India at a depth of 10 cm in May 2007 and placed in new polythene bags, using sterile spatula, prior to laboratory analysis. Starch, casein, yeast extract, peptone, l-asparagine, soybean meal, beef extract, acrylamide, bisacrylamide, bovine serum albumin and commassie brilliant blue G-250 were purchased from Hi-media (Mumbai, India). Sephadex G-100 was obtained from Sigma-Aldrich, Germany, while trichloroacetic acid was purchased from E. Merck Ltd, Mumbai, India. All other chemicals were of analytical grade and used without further modification. Enrichment and isolation of marine microorganisms One gram of sediment was transferred to a conical flask containing 100 ml of sterile seawater complex broth9, sterile starch casein broth and glucose asparagine agar prepared in natural sea water10 for the pre-enrichment of samples. The flask was incubated at 30 ºC for 14 days in a shaker incubator. A loopful of the inoculum from the pre-enriched seawater complex broth, starch casein broth or glucose asparagine broth was streaked on seawater complex agar (SWC), starch casein agar (SCA) and glucose asparagine agar, and the plates were incubated at 30 ºC for 7 days. Single discrete colonies were isolated and used for identification. Characterisation of marine soil isolates Isolated colonies were identified using standard International Streptomyces Project (ISP) procedure10. Morphological identification of isolated colonies was carried out by simple staining, Grams staining and motility testing by hanging drop method11. Biochemical characterisation was by melanoid production test using Waksman medium at a incubation temperature of 37 °C for 4 days for the detection of pigment producing property of isolates; organic nitrate reduction test was carried out in organic nitrate broth at an incubation temperature of 37 °C for one week for the detection of nitrate reducing property of isolates; acid production test was carried out in glucose nutrient broth at an incubation temperature of 20 °C for 15 days for the detection of glucose fermentation leading to the production of acid; hydrogen sulphide production test was carried out in SIM Agar at an incubation temperature of 37 °C for 5 days, while gelatin liquefaction test was carried out in nutrient gelatin at an incubation temperature of 37 °C for 24 – 48 hours for the detection of gelatin hydrolyzing properties of isolates12. Screening of soil isolates for L-asparaginase production by rapid-plate assay The isolates were screened for asparaginase activity using the method of Gulati et al13. The medium used were modified M-9 media incorporated with a pH indicator (phenol red). L-asparaginase activity was identified by formation of a pink zone around colonies. Two control plates were also prepared using modified M-9 media - one was without dye while the other was without asparagine. Crude enzyme production The crude enzyme was prepared by the methods of solid-state fermentation (SSF) and submerged fermentation (SF) Solid-state fermentation (SSF) method The fermentation media consisted of 5 g of soybean meal and 10 ml of 0.1M sodium phosphate buffer (pH 7.0) in a 250 ml flask. The flasks were autoclaved for 15 min at 121 ºC and 15 lb pressure, then cooled and inoculated with 3 ml of the previously prepared bacterial suspension. It was incubated at 37 ºC for 4 days without shaking and the crude enzyme was precipitated at the end of fermentation by the addition of 90 ml of 0.1M sodium phosphate buffer (pH 7.0) followed by centrifugation at 8000 rpm for 20 min 8. Submerged fermentation (SF) method An amount (100ml) of Tryptone Glucose Yeast extract (TGY) broth (production media, pH 7.0) comprising of glucose, 0.1g; K2HPO4, 0.1 g; yeast extract, 0.5 g; tryptone, 0.5 g; water, to 100 ml, and contained in a 250 ml Erlenmeyer flask, was inoculated separately with the soil isolates and incubated at – 28 ºC in a shaker-incubator oscillating at 200 rev/min for 24 h. At the end of the fermentation period, the crude enzyme was prepared by centrifugation at 10000 rpm for 20 min. The cell-free supernatant was taken as the crude enzyme14. Similarly,100ml Tryptone Fructose Yeast extract (TFY) broth (production media, pH 7.0) comprised of tryptone, 0.5 g; yeast extract, 0.5 g; fructose, 0.1 g; potassium dihydrogen phosphate, 0.1g; and water, to 100 ml was also inoculated with the soil isolates and incubated at – 28 ºC in a shaker-incubator oscillating at 200 rev/min for 24 h. The crude enzyme was prepared by centrifugation at 10000 rpm for 20 min15. Protein determination Protein content was determined according to the method of Lowry et al16. A stock solution of standard protein, bovine serum albumin (BSA), at a concentration of 1000 mg/ml was made. From this solution, 0.2 to 1 ml of working standard solution at concentration of 100 mg/ml was taken in test tubes. The volume was made up to 1 ml with distilled water to give concentrations ranging from 20 to 100 mg/ml. 1 ml of folins cocatteau reagent was added to each test tube. After 30 min of incubation, the absorbance was measured at 660 nm using a double beam UV – visible spectrophotometer (model SL 164, Elico, Hyderabad, India), and the protein content determined.. Determination of L-asparaginase activity The method used was essentially that of Mashburn and Wriston17. In this assay, the rate of hydrolysis of L-asparagine was determined by measuring the ammonia released using Nessler’s reaction. A mixture of 0.1 ml of enzyme extract, 0.2 ml of 0.05M Tris-HCl buffer (pH 8.6), and 1.7 ml of 0.01M L-asparagine was incubated for 10 min at 37 ºC. The reaction was stopped by the addition of 0.5 ml of 1.5M trichloroacetic acid. After centrifugation at 10000 rpm, 0.5 ml of the supernatant was diluted to 7 ml with distilled water and treated with 1 ml of Nessler’s reagent. The color reaction was allowed to develop for 10 min and the absorbance read at 480 nm with a spectrophotometer. The ammonia liberated was extrapolated from a curve derived with ammonium sulphate. One unit (IU) of L-asparaginase was defined as that amount of enzyme which liberates 1 µ mole of ammonia per minute under the assay conditions18. Purification of L-asparaginase from marine actinomycetes L-asparaginase was partially purified using the following procedures: The crude enzyme was brought to 45 % saturation with ammonium sulphate at pH 8.4 and kept overnight in a cold room at 4 ºC. It was thereafter subjected to centrifugation at 8000 rpm for 10 min at 4 ºC. The precipitate was discarded, while the supernatant was brought to 85 % saturation with ammonium sulphate and centrifuged at 8000 rpm at 4 ºC for 10 min. The precipitate from this step was collected and stored at 4 ºC 6. Dialysis tubes, which were previously soaked in 1M Tris HCl buffer, were used for the dialysis of the precipitate. The precipitate was dissolved in 1M Tris HCl buffer and dialysed against the same buffer 6. Subsequently, the dialysed samples were loaded on to a pre-equilibrated column packed with 0.05M Tris HCl Sephadex G 100. It was eluted with 0.05M Tris HCl (pH 8.4) buffer containing 0.1M KCl. A total of 30 fractions were collected at the flow rate of 5 ml/30 min. Fractions showing high activity were pooled and used for further studies6. Characterisation of partially purified L-asparaginase Effect of pH, temperature and metal ions The pH activity profile of the partially purified enzyme (pH range 3.5 to 10.5) was studied. Acetate buffer (0.1 M, pH range 3.5 -5.5), 0.1 M phosphate buffer (pH range 6.0-8.0) and 0.1M Tris HCl buffer (pH range 8.5-10.5) were used for this purpose19. The assay was carried out using 0.1 ml of appropriately diluted enzyme. Specific enzyme activity was calculated by measuring the ammonia released, based on Nessler’s reaction, by the method of Mashburn and Wriston20 described above for the determination of L- asparaginase activity.Similarly, the activity of the enzyme was measured at different temperatures ranging from 30°C to 100 °C, and at the optimum pH using the assay procedure described above20. In order to determine enzyme activity in the presence of different metal ions (Mg2+, Cu2+, Zn2+ and EDTA), different salt solutions - magnesium sulphate, copper sulphate, zinc sulphate and ethylene diamine tetraacetic acid (EDTA) - were added to enzyme substrate reaction mixture to yield final concentrations of 1, 3 and 10mM, respectively21. The specific activity of each sample was then determined by the method of Mashburn and Wriston20. Determination of Km and Vmax This is one of the important parameters for evaluating the potential usefulness of the enzyme for anti-leukaemic therapy. The initial velocity of the sample was estimated in a range of L-asparagine concentrations (4 to 160 µM). Km and Vmax were calculated from Lineweaver-Burk plot18. RESULTS Enrichment and isolation of marine actinomycetes in different media From the 20 marine sediment samples tested, only 10 sediments showed growth in enrichment media. These 10 isolates were subcultured and used for further studies. Identification of actinomycetes All the isolates produced grey and white colonies without pigmentation and showed rapid growth within two days. They were identified as Streptomyces spp. by slide culture, morphological characteristics, physiological and enzymatic properties, carbon utilization and nitrogen utilization. The samples of Streptomyces spp isolated from Tamilnadu were designated S3, S4 and S6, while those from Kerala were designated K4 and K8. The biochemical characteristics of the soil isolates are summarized in Table 1. Screening of L-asparaginase positive cultures Of the isolated Streptomyces spp. screened for L-asparaginase activity, only S3 and S4 from Tamilnadu and K8 from Kerala showed positive results in rapid plate assay method. Production of L-asparaginase All three isolates examined synthesized asparaginase with yields ranging from 24.61 to 49.23 U/ml. However, soil isolate S3 was selected for further studies because of its high productivity (49.23 U/ml). Protein concentration The concentration of protein obtained from isolated organism S3 was found to be 65 µg/ml. Table 1: Biochemical characterization of the isolated organism. (Note: ‘+’ indicates positive, and ‘-‘ indicates negative.
Table 2: Purification steps of isolated L-asparaginase
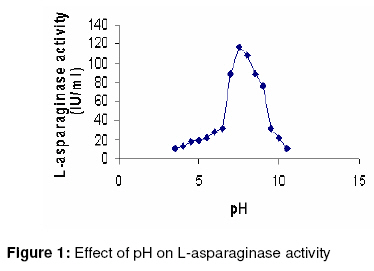
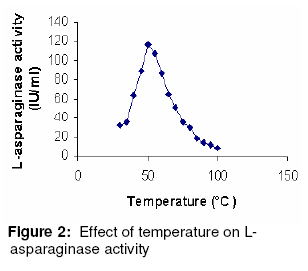
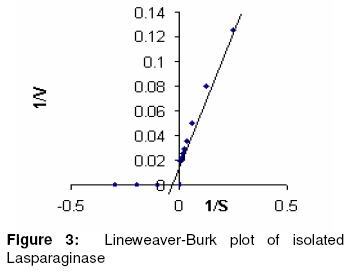
Purification and characterization of L-asparaginase The various purification steps are summarized in Table 2. In the final step, the enzyme had a specific activity of 662.61 IU/mg and approximately 2-fold purity. Figure 1 shows that the enzyme exhibited maximum activity between pH 7.0 and 8.0 with 80% activity at physiological pH. The optimum temperature was 50 ºC, as Figure 2 shows. Furthermore, the isolated enzyme was activated by Mg2+, but inhibited by Cu2+, Zn2+ and EDTA. Determination of Km and Vmax The Km and Vmax of partially purified enzyme were approximately 24 µM and 51 IU/ml, respectively, as Figure 3 shows. DISCUSSION Although L-asparaginase from bacteria has been extensively characterized, a similar attention has not been paid to actinomycetes23. To date, antibiotics are the major bioactive compounds obtained from actinomycetes. However, the ability to produce a variety of enzymes may attract research interest in these prokaryotes. Soil isolates S3 and S4 from Tamilnadu and soil isolate K8 from Kerala possess L-asparaginase activity. However the organism did not produce any pink zone in both control groups - M-9 media incorporated with L-asparagine without phenol red and M-9 media incorporated without L-asparagine. This indicates that the formation of pink zone is due only to L-asparaginase production. Among the three media used, the solid-state media gave the highest enzyme activity for all the 3 isolates. The mechanism of depressive effect in TGY and TFY broths is thought to result from the presence of glucose metabolic products. Studies suggest that in the case of asparaginase biosynthesis, the depressive effect of carbohydrates may be a function of their ability to lower the pH value of the fermentation media24. A comparison of submerged and solid-state fermentation shows a significant difference in enzyme activity. This suggests that there may be increased accumulation of intermediate metabolites between substrate and product formation in submerged fermentation. This is also probably due to the difference in the physiological state of the microorganism in solid-state and submerged fermentations. To the best of our knowledge, this is the first report on the production and partial purification of L-asparaginase from marine actinomycetes isolated via solid state fermentation (SSF). In the final purification step, the enzyme showed a specific activity of 662.61 IU/mg, which is approximately 2-fold purity. Optimum pH was found to be 7.5, which is close to blood pH, compared to L-asparaginases from other bacterial sources such as Serratia marcescens, Mycobacterium spp. and Pseudomonas spp showed optimum pH in the range of 8.0 to 8.5 15, 20, 27. At 50 °C, the enzyme showed its optimum activity. The test results of the sensitivity of L-asparaginase to heavy metal ions indicate that the activity of the enzyme may depend on the presence of sulfhydryl functional groups. The affinity of L-asparaginase to its substrate is related to its degree of effectiveness against tumors25. The substrate affinity of L-asparaginase as measured by Km value was found to be 2.4 x 10-5 M for L-asparagine. This compares favorably with Km values reported for antineoplastic L-asparaginases from other sources such as E. coli (1.25 x 10-5 M) 26, Proteus vulgaris (2.6 x 10-5 M) 27 and Erwinia aroideae NRRL Q-138 (3.0 x 10-5 M) 27. The linearity of the Lineweaver-Burk double reciprocal plot was suggests that the isolated L-asparaginase followed the Michaelis-Menten kinetics. CONCLUSION The present study only indicates that marine sediment samples from Tamilnadu and Kerala in India are potential sources of bioactive actinomycetes but also demonstrates that different pretreatment methods may be successfully applied for the isolation of actinomycetes. Hyperproductive marine actinomycetes with higher L-asparaginase activity was successfully isolated using a low-cost substrate - soyabean meal. Furthermore, isolated L-asparaginase appears to be a promising agent and requires further investigation of its potential anti-leukaemic activity. On the basis of the data obtained, marine actinomycetes may well be an unexploited source of potentially valuable products and may also be preferable to mass screening of common bacteria. Consequently, we suggest that enzymes which degrade amino acids should receive greater attention as potential therapeutic agents. REFERENCES
© Pharmacotherapy Group, Faculty of Pharmacy, University of Benin, Benin City, 300001 Nigeria. The following images related to this document are available:Photo images[pr09046f1.jpg] [pr09046f2.jpg] [pr09046f3.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}