 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Tropical Journal of Pharmaceutical Research, Vol. 9, No. 6, November-December, 2010, pp. 565-572 Research Article PFCRT and DHFR-TS Sequences for Monitoring Drug Resistance in Adzopé Area of Côte d’Ivoire After the Withdrawal of Chloroquine and Pyrimethamine L Ouattara1, KB Bla1, SB Assi2, W Yavo1,2 and AJ Djaman1,3* 1UFR Biosciences et UFR des Sciences Pharmaceutiques & Biologiques, Université de
Cocody, BP V34 Abidjan, *Corresponding author: E-mail: djamanj@yahoo.fr; Tel: +225-22 50 35 60 Received: 5 May 2010 Revised accepted: 11 September 2010 Code Number: pr10067 Abstract Purpose: Drug resistance is probably the greatest challenge to most malaria-control
programmes. The goal of this study was to evaluate polymorphisms in parasite
resistance gene markers, pfcft and dhfr, from falciparum malaria isolates collected
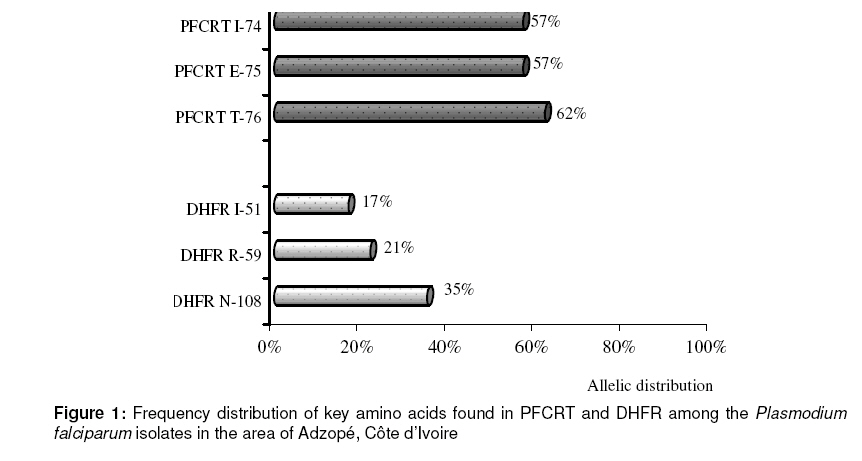
in Adzopé City, Côte d’Ivoire in 2007. Keywords: Côte d’Ivoire, Plasmodium falciparum, Malaria resistance, Molecular markers INTRODUCTION Malaria remains a major cause of morbidity and mortality in sub-Saharan Africa [1]. Worldwide malaria morbidity is estimated to be 300-500 million cases with about 1 million deaths each year, of which 90% occur in sub-Saharan Africa [2]. The vast majority of deaths occur among children below five years of age and pregnant women, especially in remote rural areas with poor access to health services [3]. Malaria control is hampered by increasing spread of drug resistance. Although sulphadoxine-pyrimethamine (SP) has been abandoned as first-or second-line treatments, respectively, by most African malaria endemic countries in favour of artemisinin-based combination treatments (ACT), it is still used as intermittent preventive treatment during pregnancy (IPTp) [4]. However, SP resistance has been on the rise in the past few years and, although the link between molecular markers and treatment failure has not been firmly established, at least for pregnant women, these markers should be monitored. Pyrimethamine inhibits dihydrofolate reductase (DHFR), whereas sulfadoxine inhibits dihydropteroate synthetase (DHPS) [5]. Resistance to these drugs has been associated with single nucleotide polymorphisms (SNP) in the genes encoding DHFR and DHPS [6]. The key amino acid substitutions that confer falciparum malaria resistance to sulfadoxine and pyrimethamine are Ala437Gly and Lys540Glu or Ala437Gly in dihydropteroate synthase (DHPS), and Ser108Asn or Ser108Thr in dihydrofolate reductase (DHFR). In addition, dhfr mutations from Asn-51 to Ile-51 and Cys-59 to Arg-59 in association with Asn-108 were found to be associated with greater resistance to pyrimethamine [7]. In most countries, chloroquine (CQ) was, for many years, the first-line drug for malaria treatment. Unfortunately, resistance to this drug is nowadays widespread. However, in areas where drug pressure has been removed, the molecular markers linked to CQ drug resistance seem to disappear, indicating that local parasites are again sensitive to this drug [8,9]. It is, therefore, important to use molecular markers for monitoring drug resistance in a country and understand its spread as this may be important in designing strategies to guide the selection of drug resistant parasites. The spread of point mutations linked to drug resistance could be monitored by carrying out regular cross sectional surveys and then genotyping the samples from infected people. Molecular tools for the detection and identification of these SNPs, such as restriction fragment length polymorphism PCR (RFLP-PCR), are available [10]. CQ resistance is associated with a mutation in the Plasmodium falciparum chloroquine resistance transporter gene (K76T) [11]. This mutation is generally not isolated but is associated, depending on the geographical setting, with mutations at other codons, Cys72Ser, Met74Ile, Asn75Glu, Ala220Ser, Gln271Glu, Asn326Ser, Ile356Thr and Arg371Ile, the role of which is not well defined. In Côte d'Ivoire, until 2003, CQ and SP were used as first-and second-line drugs, respectively, for the treatment of uncomplicated malaria. ACT replaced CQ as the recommended first-line drug. SP remains recommended for IPT among pregnant women. A few years after, it was considered important to measure the prevalence of molecular markers of chloroquine and pyrimethamine resistance. Indeed, in Malawi, a sub-Saharan country, from 1992 to 2000, the prevalence of the CQ marker gradually decreased, disappearing completely by 2001 after the withdrawal of CQ [12]. The study aimed to evaluate SP and CQ resistance in falciparum malaria isolates, following withdrawal of these drugs, in Adzopé, Côte d'Ivoire, using molecular characterization of the dhfr-ts and pfcrt genes, respectively. This city is considered as one of the sentinel sites of the national programme for malaria control. EXPERIMENTAL Blood collection area This study is a part of the cross-sectional surveys carried out in 2007 in Adzopé (lat. 06°06´N and long. 03°51´W), situated in the south of Côte d'Ivoire, 108 km from Abidjan, the economic capital city. The rainy season occurs from March to July (average rainfall: 1,789 mm/yr). The population was estimated at 43,821 inhabitants in 1998. Malaria is hyperendemic with no seasonal transmission. The most common vectors are Anopheles gambiae and Culex Quinquefasciatus. Plasmodium falciparum is the predominant malaria parasite. Blood samples used in this study were collected at random from 72 children aged between 6 months and 14 years, during a cross sectional survey in Adzopé. Fingerprick samples of capillary blood were collected from the selected children onto a filter paper strip (Isocode Stix®; Schleichler and Schuell, Ecquevilly, France). The samples were dried and stored at room temperature in an individual zip-lock polyethylene bags prior to DNA extraction and PCR studies. Plasmodium DNA extraction and amplificationDNA was extracted from the dried blood on each filter-paper strip by rinsing each strip with 500 µL distilled water, immersing it in 75 µL distilled water in a 0.5mL microtube and then incubating at 99 °C for 30 min [13]. The following mixture was prepared to a final volume of 50 µL: genomic DNA extracted as described above, 10 picomoles of each specific primer (obtained from Sigma-Aldrich®), buffer (10 mM Tris, pH 8.3, 50 mM KCl, 1.5 mM MgCl2), 0.2 mM of deoxynucleoside triphosphates (mixture of dGTP, dATP, dTTP, and dCTP), and one unit of taq DNA polymerase (Roche Diagnostics, Meylan, France). For the primary PCR, the primers pairs used were STIL (5'ATG-ATG-GAA-CAA-GTC-TGC-GAC-GTT-TTC-GAT)/ST2L(5'TTC-ATT-TAA-CAT-TTT-ATT-ATT-CGT-TTT-CTT) for dhfr-ts and TCRP-1 (5'CCG-TTA-ATA-ATA-AAT-ACA-CGC-AG) /TCRP-2 (5'CGG-ATG-TTA-CAA-AAC-TAT-AGT-TAC-C) for pfcrt. Due to the small quantity of DNA imbibed onto filter paper, nested PCR was necessary to obtain sufficient amount of amplified products for direct PCR sequencing or PCR-RFLP. So, the first amplification was performed as described above, using 12.5 µL of DNA suspension. The secondary amplification was performed on the primary amplification product with internal primers: ST1L/DHFR-595R (5'CTG-GAA-AAA-ATA-CAT-CAC-ATT-CAT-ATG-TAC) for dhfr-ts (595bp fragment) and TCDR-1 (5'TGT-GCT-CAT-GTG-TTT-AAA-CTT) /TCDR-2 (5'CAA-AAC-TAT-AGT-TAC-CAA-TTT-TG) for pfcrt (145 pb fragment). For the all running PCRs, the PTC-100 thermal cycler (MJ Research, Watertown, MA, USA) was programmed to carry out 30 cycles of 94 °C for 2 min (first cycle) or 1 min (rest of the cycle), 50°C x 1 min (56 x 30 s for pfcrt), and 72 °C x 1min, followed by 72 °C x 10 min at the end of 30 cycles. PCR-RFLP and sequencing of DNA fragmentsPfcrt PCR products were digested by Apo I at 50°C in the presence of NE buffer3 (50mM Tris-HCl, 10mM MgCl2, 100mM NaCl, 1mM DDT). As for dhfr-ts PCR products, they were submitted to the digestion of Alu I {(37 °C, NE buffer 2 (10mM Tris-HCl, 10mM MgCl2, 50mM NaCl and 1mM DTT)}. Apo I and Alu I cut the wild DNA fragments, haplotype Lys76 and Ser-108 of pfcrt and dhfr-ts genes, respectively. However, in spite of the use of these enzymes, all the PCR products or DNA fragments from agarose gel (mixed alleles) were purified using the High Pure Kit purification product (Roche Diagnosis) and then sequenced in order to investigate additional mutations. Quality control and Data analysisA known DNA of P. falciparum was used as positive control in the PCR evaluation. The electropherograms of the purification PCR products sequenced by MWG-biotech (MWG-Eurofin Operon) were analyzed by using 4 peaks, Chromas Lite version 2.01 and ApE-A plasmid Editor V1.12 software. According to the haplotypes 74, 75 and 76 of pfcrt gene investigated, the wild-type pfcrt isolate was defined by the following combination: Met74/Asn75/Lys76, while for dhfr-ts, the allelic combination Ala16/Asn51/Cys59/Ser108/I164 defined the wild-type isolates. However, for each gene, the key amino-acid mutations that confer the resistance are Lys76 for pfcrt and Ser108 for dhfr-ts, respectively. Ethical reviewThe study protocol was approved by the ethics committee of the Ivorian Ministry of Public Health and all legal parents or guardians of enrolled children gave their consent for blood sampling. RESULTSA total of 72 samples collected were successfully amplified. After PCR-RFLP running, according to only the haplotype 76 of pfcrt, 21 (29 %) wild type pfcrt Lys76, 41 (57 %) mutant type pfcrt Thr76 and 10 (14 %), mixed population wild type/mutant type DNA fragments were obtained. For dhfr-ts, the outcome confirmed again the presence of 10 wild type/mutant type (Ser108Asn) dhfr-ts isolates against 19 mutant-type dhfr-ts Asn108 (Table 1). Due to the presence of the 10 mixed alleles, a total of 82 DNA fragments were sequenced instead of 72. Among them, 31 (37.8 %) were wild-type pfcrt (Met74/Asn75/Lys76) against 51 (62.2 %) mutant type pfcrt. The mutant pfcrt haplotypes consisted of four single mutations (Met74/Asn75/Thr76) and 47 triples mutations (Ile74/Glu75/Thr76). No double mutations were found (Table 1). The distribution of each PFCRT allele was: 57.3 % (47) for both Ile74 Glu75, while 51 (62.2 %) samples of DNA fragments were Thr76 mutations (Table 2, Figure 1). Of the 82 samples of P. falciparum isolates investigated, 53 (64.6 %) had wild-type dhfrts, while 29 (35.4 %) were mutant-type dhfrts. They consisted of four single mutant-type samples (Asn51/Cys59/Asn108), nineteen double mutations distributed as follows: Ile51/Cys59/Asn108 (eight) and Asn51/Arg59/Asn108 (eleven). Six samples were triple mutant-type dhfr-ts Ile51/Arg59/Asn108 mutation (Table 2). DISCUSSION Drug resistance is probably the greatest challenge to most malaria control programmes. Given the limited resources for other malaria control measures, rational drug use is crucial, although economic constraints and the scarcity of the drug of choice hampers this. Close monitoring of the epidemiology and dynamics of drug resistance are necessary to implement measures to circumvent the problem and to allow Côte d’Ivoire’s National Malaria Control Programme to recommend the best available management of malaria. Like the in vitro tests, molecular markers of resistance might provide an early warning system in geographical or temporal monitoring for guiding treatment efficacy studies [14]. Molecular markers are useful for monitoring the prevalence of mutations after a drug has been withdrawn or when a drug combination is used [12]. Our study was focused on molecular markers for chloroquine and pyrimethamine resistance. This study reveals a high frequency of drug resistance molecular markers for CQ (pfcrt Thr76; 62.2%). A similar study carried out in Abidjan, an area near Adzopé, reported 65 % of mutant type pfcrt Thr76 [15]. Previous studies conducted in Côte d’Ivoire and elsewhere in West Africa showed strong relationship between pfcrt Thr76 and therapeutic failure or in vitro chemoresistance of Plasmodium falciparum isolates to chloroquine [16,17]. This mutation is not generally isolated but is associated with other mutations at haplotypes 72, 74, 75, etc. In this study, most of the mutant-type pfcrt were triple mutant (Ile74/Glu75/Thr76) but the role of these complementary mutations is not well defined. A few years later, chloroquine was withdrawn from use due to high treatment failure rates (> 44 %). The proportion of pfcrt mutation (62 %) is reasonable evidence that there is a high level of chloroquine-resistant parasites circulating in this area. Thus, it would be too early to consider returning to the use of chloroquine alone as an antimalarial, although the drug itself is a safe and inexpensive, and ideally may be used in combination with another suitable antimalarial drug. In contrast, in Malawi, the first African country to replace chloroquine with SP, from 1992 to 2000, the prevalence of the molecular marker of chloroquine gradually decreased, disappearing completely by 2001 [12]. Today, chloroquine is once again highly efficacious in Malawi, 12 years after cessation of chloroquine use in that country [8]. For dhfr-ts gene, the mutation mainly affected codons 108 (35 %), 59 (21 %) and 51(17 %). Quadruple mutant, implying also the haplotype 16 or 164 which represents the most severe form of resistance and is responsible for high level of resistance to pyrimethamine, is at the present time not among the isolates of Côte d’Ivoire. These results are in agreement with others from western Africa, where prevalence of quadruple mutant has been observed [18]. A few rare strains carry the quadruple mutations in Africa, namely, Ghana, Kenya and Tanzania [19,20]. Elsewhere in the world, the Ile164Leu mutation is relatively uncommon but has been reported in Bangladesh, Bolivia, Brazil, Cambodia, China, India, Malaysia, Peru, Thailand and Viet Nam [21,22]. Although the frequency of mutation of dhfr-ts (Asn108, 35.4%) is relatively less than that seen with CQ in the same area (62.2 %), it is still alarming considering that the use of sulphadoxine-pyrimethamine as intermittent preventive treatment in pregnancy is unavoidable because suitable alternatives are not available yet. However, the prevalence of molecular markers of malarial organisms resistant to both chloroquine and pyrimethamine represented 48.8 % of the total samples. One of the reasons for this high level of the molecular markers of CQ and SP resistance is probably the wide usage of these drugs in Côte d’Ivoire in spite of their official replacement with ACT for the treatment of uncomplicated falciparum malaria. Maybe it is too early to evaluate the sensitivity of these drugs by monitoring the molecular markers for falciparum malaria resistance. In that case, it would be necessary to wait for some years before embarking on another study of this type. CONCLUSIONThe study was the first molecular study carried out in this geographical area which is considered as one of the sentinel sites for malaria control programme of Côte d’ivoire. The results of the epidemiological study on the prevalence of genotypes associated with drug resistance, carried out in Adzopé, showed a high presence of chloroquine resistance markers, while for SP a lower prevalence was detected. Since 2003, following the withdrawal of these antimalarials as first-and second-line drugs, respectively, the status of chloroquine and pyriméthamine-resistant parasite has not changed much in Adzopé with regard to the mutations affecting dhfr-ts and pfcrt genes. ACKNOWLEDGEMENTThe authors are very grateful to Drs M Adja, AL Ahou and NB Tchiekoi for assisting in the collection of P. falciparum samples in the study area. Thanks are also due to Dr A Mazabraud (Director of Research) who provided facilities for the PCR test in his laboratory (CNRS-UPR 3294 Bat.445, Université Paris Sud, 91405 Orsay Cedex, France). REFERENCES
Copyright 2010 - Tropical Journal of Pharmaceutical Research The following images related to this document are available:Photo images[pr10068t2.jpg] [pr10068t1.jpg] [pr10068f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}