 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
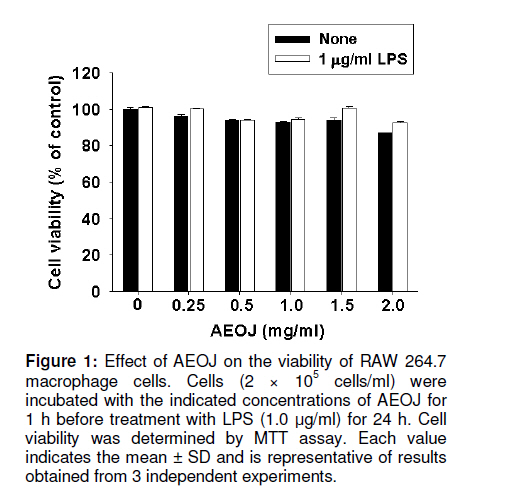
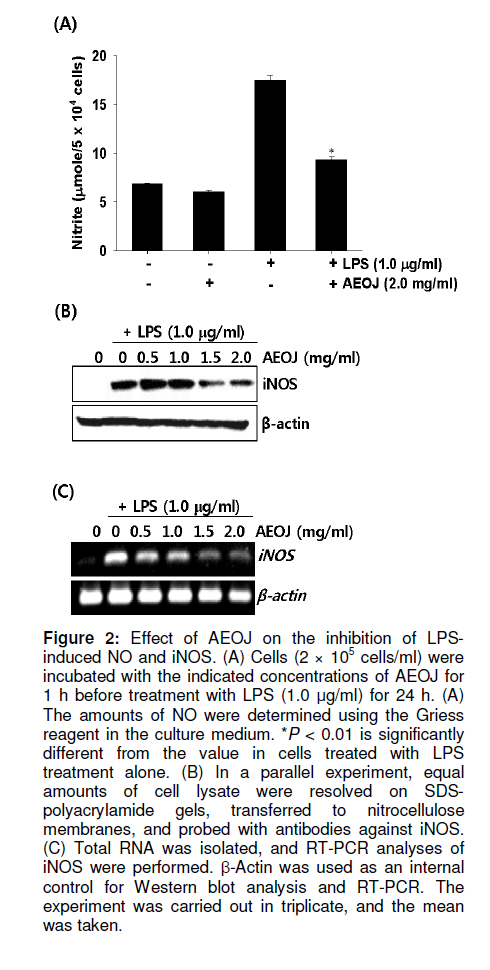
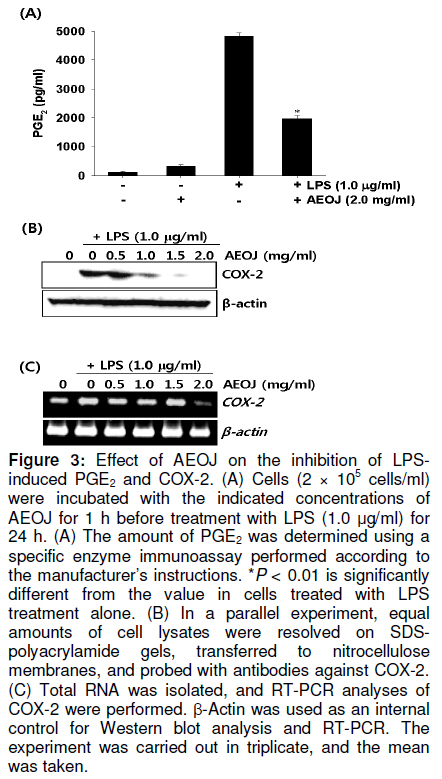
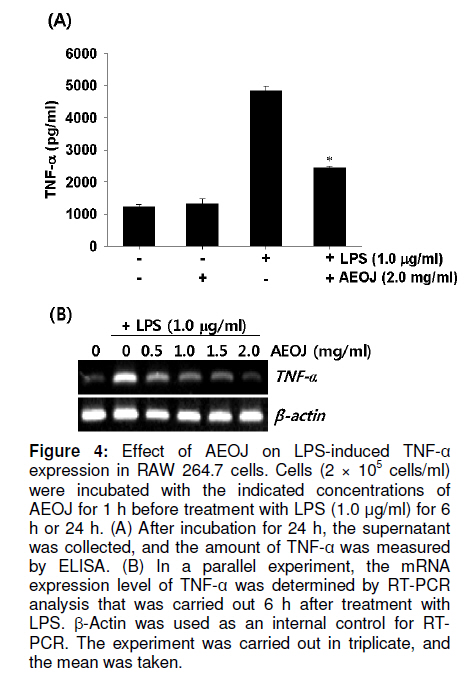
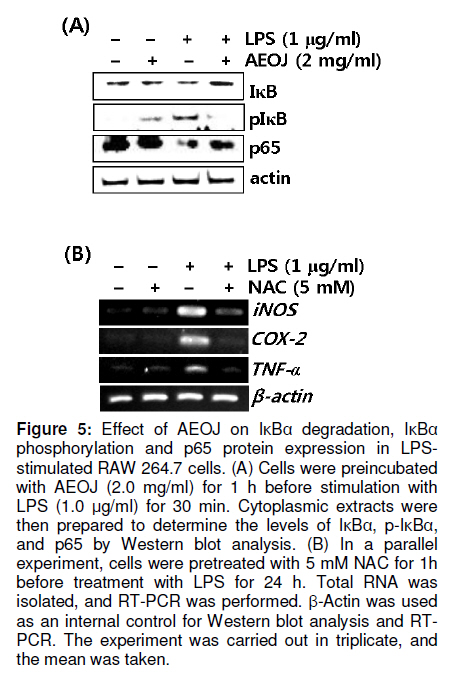
Tropical Journal of Pharmaceutical Research, Vol. 10, No. 2, April, 2011, pp. 161-168 Research Article Inhibition of Lipopolysaccharide-Induced iNOS, COX-2, and TNF-α Expression by Aqueous Extract of Orixa Japonica in RAW 264.7 Cells via Suppression of NFκB Activity Chang-Hee Kang1, Yung H Choi2, Il-Whan Choi3, Jae-Dong Lee4 and Gi-Young Kim1,* 1Laboratory of Immunobiology, Department of Marine Life Sciences, Jeju National University, Jeju 690-756, Received: 16 September 2010 Revised accepted: 19 January 2011 Code Number: pr11023 Abstract Purpose: To investigate the anti-inflammatory effects of aqueous extract of Orixa japonica (AEOJ) in lipopolysaccharide (LPS)-stimulated RAW 264.7 cells. Keywords: Orixa japonica, Nitric oxide, Prostaglandin E2 , Tumor necrosis factor-α, Nuclear factor-κB INTRODUCTION Orixa japonica is a plant that is widely distributed in East Asia. Its aqueous extract AEOJ has been used traditionally for the treatment of cough, arthritis, dysentery, and malaria [1-3]. Sharma and his colleagues also reported that AEOJ is a good pharmaceutical candidate for the treatment of obesity caused by high fat-diet [4]. More recently, it was found that AEOJ significantly inhibits the growth of acne-inducing pathogenic bacteria, is not cytotoxic to the human cell line, and suppresses acne-inducing inflammatory cytokines in THP-1 cells [5]. However, little is known about the mechanism by which AEOJ inhibits lipopolysaccharide (LPS)-induced inflammation. Inflammation is typified by the activation of immunocytes such as monocytes and macrophages, and the secretion of inflammatory mediators such as nitric oxide (NO), prostaglandin E2 (PGE2), and tumor necrosis factor-α (TNF-α). Although NO plays a significant role in host immune defense, vascular regulation, neurotransmission, and other systems under normal conditions, aberrant NO expression is thought to cause severe inflammatory disease [6,7]. Overproduction of inducible NO synthase (iNOS) is especially related to various human diseases such as inflammatory and neuronal disorders because of the upregulation of NO [8,9]. PGE2 , which is derived from cyclooxygenase-2 (COX-2) by inflammatory stimulants, is also thought to be involved in the pathogenesis of some inflammatory diseases [10,11]. Additionally, TNF-α is highly expressed in macrophages involved in the inflammatory process and causes severe tissue damage, septic shock, atherosclerosis, and systemic inflammatory response syndrome [12,13]. Therefore, the inhibition of these inflammatory mediators would be an effective therapeutic approach for regulating LPS-induced septic shock. NF-κB is a nuclear transcription factor that regulates various genes related to immune and inflammatory responses, cell proliferation, growth, cell adhesion, and survival [14-15]. It is well known that NF-κB regulates the expression of inflammatory genes such as iNOS, COX-2, and TNF-α in order to boost inflammatory responses in early stages [16]. Previous studies also demonstrated that the inhibition of NF-κB activity may be useful for attenuating various inflammatory diseases including rheumatoid arthritis, septic shock and ischemia [17-19]. Therefore, effective inhibitors of NF-κB may explain its immunomodulatory and anti-inflammatory effects in LPS-induced inflammation. In the present study, an effort was made to show that AEOJ significantly inhibited the expression of iNOS, COX-2, and TNF-α, and their main products via the suppression of NF-κB activity. EXPERIMENTAL Preparation of plant extract (AEOJ) O. Japonica was purchased from a local oriental herb store, Kwang Myoung Dang (Busan, Republic of Korea) in February 2005, and authenticated by Professor WS Ko, College of Oriental Medicine, Dongeui University, Busan, Republic of Korea. A voucher specimen was deposited at the Department of Oriental Medicine, Dongeui University. The plant material (100 g) was percolated with distilled water at 100°C for 2 h. The extract was filtered through 0.45 µm filter and the filtrate freeze-dried. The extract yield was approximately 4.1g and the extract (AEOJ) was kept at 4°C until used. The extract was dissolved in phosphate buffered saline (PBS) at 1 mg/ml and filtered through 0.22 µm filter before use. Reagents LPS and 3-(4,5-dimethylthiazol-2-yl)-2,5diphenyl-tetrazolium bromide (MTT) were purchased from Sigma Chemical Co. (St. Louis, MO, USA). Rabbit anti-human antibodies against iNOS, COX-2, p65, IκBα, and phospho (p)-IκBα were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). The antibody against β-actin was obtained from Sigma. Peroxidase-labeled goat anti-rabbit immunoglobulin was purchased from KOMA Biotechnology (Seoul, Republic of Korea). Dulbecco’s modified Eagle’s medium (DMEM) and fetal bovine serum (FBS) were obtained from WelGENE Inc. (Daegu, Republic of Korea). Other chemicals were purchased from Sigma. Cell Culture and viability RAW 264.7 murine macrophages cell line was kindly obtained from SJ Jeong (Kyung Hee University, Seoul, Republic of Korea) and cultured at 37°C in 5% CO2 in DMEM medium supplemented with 10% FBS and antibiotics (WelGENE). For the analysis of cell viability, the cells were incubated with the indicated concentrations of AEOJ for 1 h prior to the treatment with LPS (1 µg/ml) for 24 h. Cell viability was determined by MTT assay. Isolation of total RNA and RT-PCR Total RNA was extracted from RAW 264.7 cells using Trizol reagent (Invitrogen, Carlsbad, USA) according to the manufacturer’s instruction. One microgram of RNA was reverse-transcribed using MMLV reverse transcriptase (Bioneer, Daejeon, Republic of Korea). Then, cDNA was amplified by PCR using specific primer iNOS (forward 5’-cct cct cca ccc tac caa gt-3’ and reverse 5’-cac cca aag tgcttc agt ca-3’), COX-2 (forward 5’-aag act tgc cag gct gaa ct-3’ and reverse 5’-ctt ctg cag tcc agg ttc aa-3’), TNF-α (forward 5’-gcg acg tgg aac tgg cag aa-3’ and reverse 5’-tcc atg ccg ttg gcc agg ag-3’) and β-actin (forward 5’-tgt gat ggt ggg aat ggg tca g-3’ and reverse 5’-ttt gat gtc acg cac gat ttc c-3’). The following PCR conditions were applied: COX-2 and iNOS, 25 cycles of denaturation at 94°C for 30 s, annealing at 59°C for 30 s and extended at 72°C for 30 s; β-actin, 23 cycles of denaturation at 94°C for 30 s, annealing at 57°C for 30 s and extended at 72°C for 30 s. β-actin was used as an internal control to evaluate relative expression of COX-2, iNOS and TNF-α. Western blot analysis Total cell extracts were prepared using PRO-PREP protein extraction solution (iNtRON Biotechnology, Sungnam, Republic of Korea). Cell lysates were centrifuged at 14,000 g at 4°C for 10 min to obtain the supernatants. Supernatants were collected and protein concentrations determined using a Bio-Rad protein assay kit (Bio-Rad, Hercules, CA, USA). Samples were stored at -80°C or immediately used for Western blot analysis. The proteins were separated on SDS-polyacrylamide gels and transferred to nitrocellulose membranes (Schleicher & Schuell, Keene, NH, USA). Proteins were detected using an enhanced chemiluminescence detection system (Amersham, Arlington Heights, IL, USA). NO Assay RAW 264.7 cells (2 × 105 cells/ml) were plated onto 24-well plates and pretreated with the indicated concentrations of AEOJ for 1 h prior to stimulation with 1 µg/ml of LPS for 24h. Briefly, the sample supernatants were mixed with equal volume of Griess reagent (1% sulfanilamide in 5% phosphoric acid and 0.1% naphthylethylenediamine dihydrochloride) and then incubated at room temperature for 10 min. The absorbance was measured at 540 nm on a microplate reader (Thermo Electron Corporation, Marietta, OH, USA). Nitrite concentration was determined using a dilution of sodium nitrite as a standard. Measurement of TNF-α and PGE2 The expression levels of TNF-α and PGE2 were measured by enzyme-linked immunosorbent assay (ELISA) kit (R&D Systems, Minneapolis, MN, USA) according to the manufacturer’s instructions. Briefly, RAW 264.7 cells (2 × 105 cells/ml) were plated in 24-well plates and pretreated with the indicated concentrations of AEOJ for 1 h prior to stimulation with 1 µg/ml of LPS for 24 h. One hundred microliters of culture-medium supernatants were collected for the determination of PGE2 and TNF-α concentrations by ELISA. Statistical analysis All data were derived from at least three independent experiments. Statistical analyses were conducted using SigmaPlot software (version 6.0) Values were presented as mean ± SD. Significant differences between the groups were determined using the unpaired Student’s t-test. Statistical significance was set at p < 0.05. RESULTS AEOJ is not cytotoxic to RAW 264.7 cells To assess the effects of AEOJ on the viability of RAW 264.7 cells, we treated cells with the indicated concentrations of AEOJ in the presence or absence of LPS for 24 h and analyzed cell viability by MTT assay. The viability of RAW 264.7 cells was not significantly altered by 24-h incubation with up to 2 mg/ml of AEOJ (Figure 1). Additionally, in the presence of LPS (1 µg/ml), AEOJ did not affect the viability of the cells. Therefore, in subsequent experiments, the concentration of AEOJ used was 2 mg/ml. AEOJ suppresses the expression of iNOS and NO To investigate whether AEOJ regulates NO production, cells were pretreated with AEOJ for 1 h before treatment with LPS for 24 h, and NO production was measured by performing the Griess reagent assay. NO production was reflected in the accumulation of nitrite in the cell culture medium. Treatment with LPS resulted in significant upregulation of nitrite production (17.3 ± 0.5 µM), compared to the untreated control (6.2 ± 0.4 µM; Figure 2A). However, RAW 264.7 cells pretreated with AEOJ displayed a marked decrease in the induction of nitrite (9.4 ± 0.3 µM) after stimulation with LPS. Next, we investigated whether AEOJ regulates iNOS expression at the protein and gene levels. Consistent with the suppression of nitrite production, AEOJ induced dose-dependent downregulation of iNOS expression at the protein (Figure 2B) and gene levels (Figure 2C). These data indicate that AEOJ attenuates the upregulation of LPS-induced NO production by suppressing iNOS at the transcriptional level. AEOJ suppresses the expression of COX-2 and PGE2 To determine whether AEOJ suppresses the production of PGE2 in LPS-stimulated RAW 264.7 cells, cells were stimulated with LPS and the amount of PGE2 secreted was measured using anti-PGE2 -coated ELISA plates. RAW 264.7 cells stimulated with LPS significantly increased the amount of PGE2 secreted (4826 ± 123 pg/ml) as compared to untreated controls (145 ± 23 pg/ml; Figure 3A); however, this induction was markedly suppressed in AEOJtreated cells (1978 ± 98 pg/ml). Next, we confirmed the expression of COX-2 in RAW 264.7 cells by Western blot analysis and RT-PCR. Treatment with AEOJ decreased LPSinduced COX-2 expression at the protein level in a dose-dependent manner (Figure 3B). Additionally, RT-PCR data showed that the mRNA expression of COX-2 was increased by LPS treatment; however, pretreatment with AEOJ reversed LPS-induced COX-2 mRNA expression in RAW 264.7 cells. These data indicate that AEOJ suppresses the upregulation of LPS-stimulated PGE2 expression via the suppression of COX-2 expression at the transcriptional level. AEOJ inhibits production of TNF-α at the gene level Next, we attempted to test the potential effects of AEOJ on the mRNA expression and secretion of TNF-α in LPS-stimulated RAW 264.7 cells. Cells were pretreated with 2 mg/ml AEOJ for 1 h before treatment with LPS for 24 h. TNF-α production was assessed in the culture supernatant by ELISA. TNF-α was slightly expressed in untreated controls (1091 ± 78 pg/ml). However, LPS stimulation significantly increased TNF-α secretion at 24 h (4826 ± 156 pg/ml; Figure 4A). Pretreatment with AEOJ significantly suppressed LPS-induced TNF-α secretion (2430 ± 57 pg/ml), even though AEOJ itself did not alter the release of TNF-α in the absence of LPS. To evaluate whether the downregulation of AEOJ-induced TNF-α release was due to the regulation of the TNFα gene in LPS-stimulated RAW 264.7 cells, we performed RT-PCR analysis at 6 h after treatment with LPS. AEOJ reduced LPS-induced expression of TNF-α mRNA in a dose-dependent manner (Figure 4B). These data indicate that AEOJ suppressed LPS-stimulated TNF-α release at the transcriptional level. AEOJ attenuates the expression of inflammatory mediator gene products via the suppression of NF-κB activity Because it is well known that NF-κB activation by LPS induces the expression of inflammatory mediators such as iNOS, COX2, and TNF-α, we performed Western blot analysis to investigate whether AEOJ regulates NF-κB activity. Because the translocation of NF-κB into the nucleus is preceded by the phosphorylation and degradation of the IκB protein, we analyzed the expression of IκB, p-IκB, and p65 in the cytosol. AEOJ inhibits LPS-induced degradation and phosphorylation of IκB (Figure 5A). In addition, AEOJ helped sustain p65 expression in the cytosol. In order to test whether NF-κB induces iNOS, COX-2, and TNF-α expression, we treated cells with a non-cytotoxic concentration of NAC, an NF-κB inhibitor, and performed RT-PCR to check the expression of inflammatory mediator gene products. NAC significantly reversed LPS-induced iNOS, COX-2, and TNF-α expression at the transcriptional level (Figure 5B). We confirmed that AEOJ was capable of attenuating LPS-induced inflammatory mediators via the inhibition of NF-κB activation. DISCUSSION Although AEOJ has been used traditionally for the treatment of cough, arthritis, dysentery and malaria in Asia [1-3], little is known about the molecular mechanisms underlying its anti-inflammatory activities. In the present study, we showed that AEOJ remarkably attenuated the expression of inflammatory gene products such as iNOS, COX-2, and TNF-α at the protein and mRNA levels through the NF-κB pathway. It is well known that NO and PGE2 are the main macrophage-derived inflammatory mediators [20-21]. Furthermore, it has been reported that TNF-α is an inflammatory cytokine, which is considered as an endogenous mediator in LPS-induced shock [22]. Aberrant production of NO, PGE2 , and TNF-α induces an inflammatory response that causes damage to neighboring cells and tissues of the host. Therefore, it has been thought a good strategy to reduce LPS-induced inflammation through the suppression of these inflammatory mediators. Among the overproduced inflammatory mediators, NO has been strongly implicated in the pathogenesis of several disease processes such as septic shock, rheumatoid arthritis, and autoimmune diabetes [23]. Furthermore, PGE2 and TNF-α are highly detectable in the serum after induction by pro-inflammatory responses, which are implied in the pathogenesis of sepsis and inflammation [24,25]. According to the results of the present study, AEOJ suppressed the production of NO, PGE2 , and TNF-α in LPS-stimulated RAW 264.7 cells via the regulation of transcriptional expression. The mechanism by which AEOJ inhibits the expression of these gene products seems to involve the regulation of the common transcription factor. NF-κB is present in the cytoplasm as an inactivated dimer composed of p65 and p50 subunits. In response to inflammatory stimuli, IκB is phosphorylated and degraded, and NF-κB is released and translocated into the nucleus [26]. Because the expression of many inflammatory genes, including iNOS, COX-2, and TNF-α, are known to be modulated by the binding of NF-κB to its specific promoter regions [27], it is a good target for suppressing NF-κB activity for the regulation of LPS-induced inflammation. Our results showed that AEOJ decreases LPS-induced phosphorylation of IκB and the resultant degradation of the IκB protein. It also helped sustain the expression of the p65 protein in the cytoplasm. Pretreatment of cells with AEOJ suppressed this signal; this finding suggests that AEOJ inhibits NF-κB activation via inhibition of IκB phosphorylation. Taken together, these results suggest that AEOJ inhibits LPS-induced NF-κB activity by suppressing the degradation and phosphorylation of IκB. However, we can-not rule out the possibility that the inflammatory response is attenuated via the inhibition of other transcription factors. CONCLUSION This study confirmed that AEOJ has anti-inflammatory effects, via the suppression of NF-κB activation in RAW 264.7 cells, thereby suppressing the release of NO, PGE2 , and TNF-α. Therefore, understanding the mechanism of AEOJ action may lead to the development of more effective therapies for LPS-induced inflammatory diseases. ACKNOWLEDGMENT This study was carried out under the auspices of a grant from Jeju National University, Jeju Republic of Korea. REFERENCES
Copyright © 2011 - Pharmacotherapy Group, Faculty of Pharmacy, University of Benin, Benin City, 300001 Nigeria. The following images related to this document are available:Photo images[pr11023f2.jpg] [pr11023f3.jpg] [pr11023f5.jpg] [pr11023f1.jpg] [pr11023f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}