 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
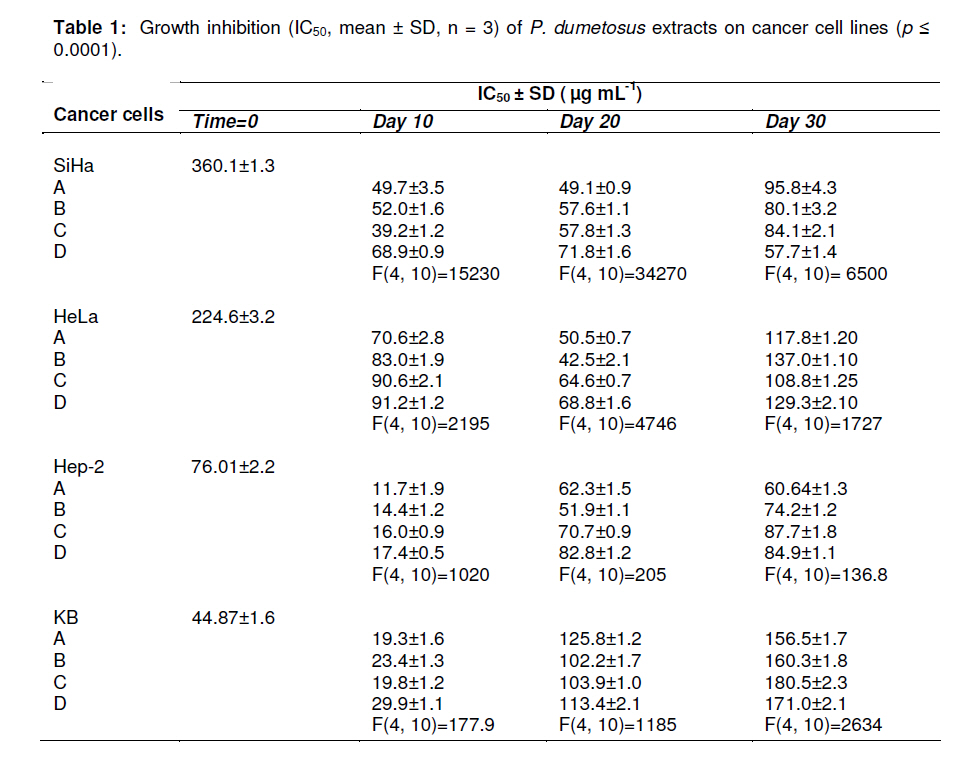
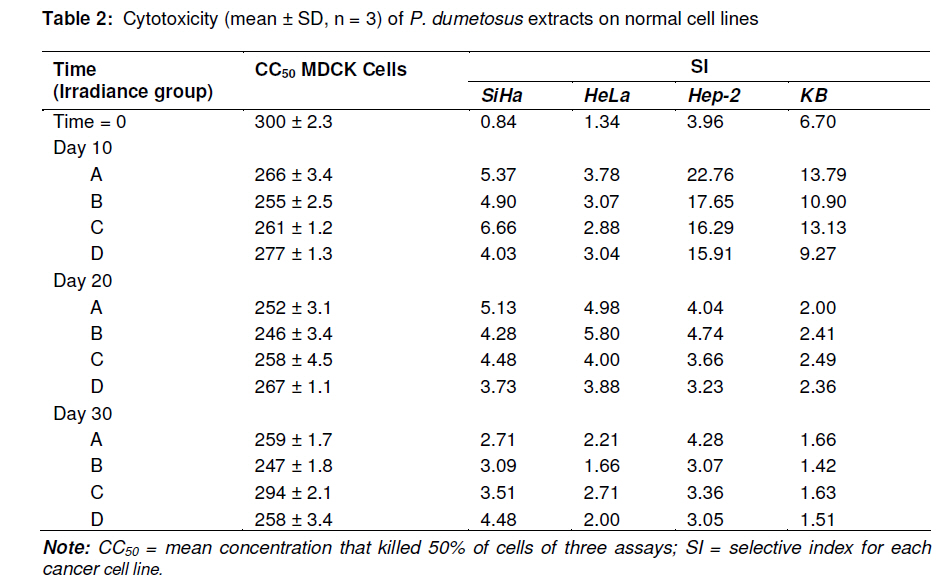
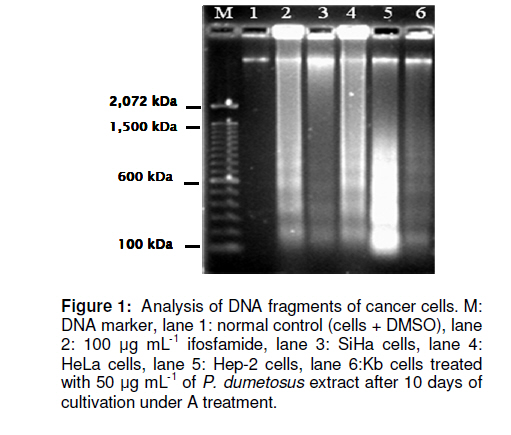
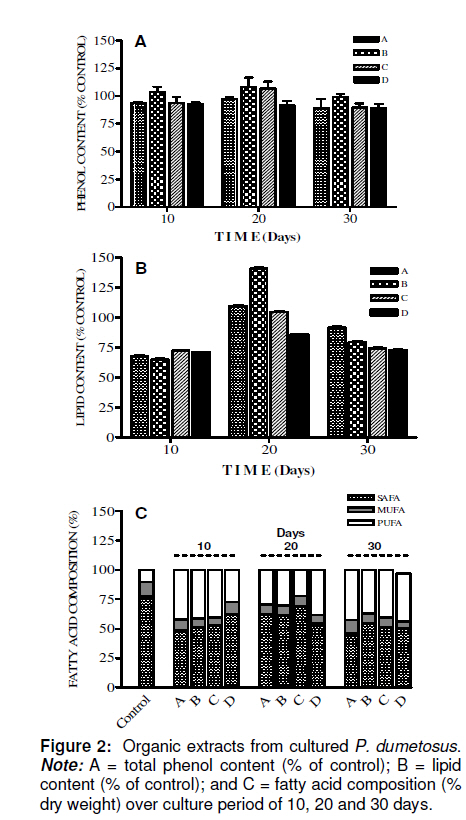
Tropical Journal of Pharmaceutical Research, Vol. 10, No. 2, April, 2011, pp. 177-185 Research Article Improved Antitumoral Activity of Extracts Derived from Cultured Penicillus dumetosus Rosa Moo-Puc1,2, Daniel Robledo1 and Yolanda Freile-Pelegrin1* 1Department of Marine Resources; Cinvestav, Km 6 Carretera Antigua a Progreso, Cordemex, 97310, A.P. 73, Mérida, Yuc., Received: 7 October 2010 Revised accepted: 31 January 2011 Code Number: pr11025 Abstract Purpose: Extracts of the green alga, Penicillus dumetosus, have shown antiproliferative activity against several cancer cell lines. The aim of this study was to evaluate if this activity is modified after cultivation. Keywords: Antiproliferative, Cancer cell lines, Bioactivity, Cytotoxicity, Green alga, Penicillus dumetosus INTRODUCTION Recent trends in drug research on natural sources suggest that algae are a promising source of novel biochemical active substances [1]. To survive in a competitive environment, marine algae have developed defense strategies that result in a significant level of structural chemical diversity that derive from different metabolic pathways [2]. The feasibility of algal cultivation can help to induce adaptations that can be measured through metabolite synthesis or biological activity. Fully controlled greenhouse-based cultivation systems have been developed for high quality year-round vegetable production for the botanical drug market [3]. Therefore, a better understanding of the potential manipulation of algal culture conditions to modify metabolite synthesis and activity is required. Tropical green alga in the order Bryopsidales, including those of the genera Avranvillaea, Caulerpa, Halimeda, Penicillus and Udotea, are noted for the production of sesqui and diterpenoids, compounds that have also shown antifungal and antiproliferative activity [2,4]. Recent studies have shown that both aqueous and organic extracts of Penicillus dumetosus exhibit in vitro antiprotozoal [5,6], as well as cytotoxic and antiproliferative activities on cancer cell lines [7]. In some cases, the antiproliferative activity of marine alga extracts has been positively correlated with the total polyphenol content suggesting a causal link related to extract content of polyphenols and phenolic acids [8], while other authors have reported a variety of fatty acids and derivatives with antiproliferative effects in different cancer cell models [9]. The high diversity of algae species coexisting in tropical habitats makes them highly competitive and complex. As a result of this intense competition, a high percentage of species have evolved chemical means of defense against predation or overgrowth by competing species [10]. Nutrient, light, water current, and temperature represent additional growth limiting components, further fueling competition. Despite the observations of antiproliferative activities in marine algae, there is limited information on how this activity may change under contrasting environmental conditions. Therefore, the objective of this study was to investigate the antiproliferative activity of crude organic extracts of cultured P. dumetosus on four human malignant cell lines (Hela, Hep-2, SiHa and KB). Furthermore, the study evaluated whether phenol content and lipid composition were related to its antiproliferative activity and to the cells’ apoptosis inducement. EXPERIMENTAL Culture conditions Penicillus dumetosus (J.V. Lamouroux) Blainville were collected in the Yucatan Peninsula coast, Yucatan, Mexico, stored in plastic bags and chilled in ice during transport to the Cinvestav Marine Station at Telchac, Yucatan, Mexico. Algae cultivation was performed under four light treatments: full (100 %) sunlight, 75 % sunlight, 50 % sunlight, and 0 % sunlight, designated A, B, C and D sections, respectively. Agricultural greenhouse shade net was used in order to obtain variable light intensities in the culture system. Light intensity varied over cultivation time: During the first 10 days, the photon flux density (PFD) in full sunlight and 75 % sunlight sections were not significantly different (one-way ANOVA, F[3,36] = 68.21, P < 0.0001; post hoc Tukey’s test P <0.0001) at 55 ± 12.9 and 65 ± 12.5 µmol photon m-2 s-1 , respectively, while 50 and 0 % sunlight sections received 60 and 3 % of incident PFD, respectively, i.e., 42 ± 12.0 and 2 ± 0.7 µmol photon m-2 s-1 , respectively. At 20-and 30-day cultivation, a similar trend was registered with 75 % sunlight section receiving 75 – 100 % and 50 % sunlight section 42 – 52 % of incident PFD; 0 % sunlight section only received 2 % of incident PFD. Preparation of extracts Freshly collected samples of the wild material were lyophilized and milled into powder to perform plant extraction protocols and analytical methods; this material was considered as control before cultivation (time 0). Entire plants of P. dumetosus (n = 10 15) were taken from each culture treatment - A, B, C and D - at 10, 20 and 30 days of the experimental period to perform organic extraction and analysis. Lyophilized samples (10 g) were exhaustively extracted with 200 mL of dichloromethane: methanol (7:3) by maceration for 24 h at room temperature. The extracts was filtered and concentrated to dryness under vacuum at 40°C and stored at -20°C until evaluated. The extracts were labeled according to culture conditions: light intensity (A, B, C or D) and time (10, 20 and 30 days). Cell line cultivation The following cell lines were used for the cytotoxicity and antiproliferative assays: normal Mardin-Darbin cell kidney (MDCK), and four human carcinoma cells, namely, laryngeal (Hep-2), cervix (HeLa), cervix squamous (SiHa) and nasopharynx (KB). The cells were grown in DMEM (Gibco) media supplemented with 10 %v/v fetal bovine serum (FBS, Gibco) with 100 U mL-1 penicillin and 100 mg/mL streptomycin. Cell lines were maintained at 37ºC in a 5% CO2 atmosphere with 95% humidity, and culture medium was changed once every 5 days. Cytotoxicity assays The cytotoxicity assay was performed according to the method of Rahman et al [11]. In this procedure, 1.5 x 104 viable cells from each cell line were seeded in a 96-well plate (Costar) and incubated for 24 to 48 h. When cells reached > 80 % confluence, the medium was replaced and the cells were treated with the organic extracts dissolved in dimethyl sulfoxide (DMSO at a maximum concentration of 0.05%) at concentrations of 6.25, 12.5, 25 and 50 µg mL-1. After 72 h of incubation, 10 µL of a 0.005% 3-(4,5dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT, Sigma) solution (5 mg mL-1) was added to each well and incubated at 37ºC for 4 h. The medium was removed and formazan, generated by the activity of dehydrogenases, was dissolved in acidified isopropanol (0.4M HCl). The amount of MTT-formazan is directly proportional to the number of living cells and was determined by measuring the optical density (OD) at 540 nm using a Bio-assay reader (BioRad, USA). Untreated cells were used as negative control. The concentration of the organic extract that killed 50% of the cells (CC50) was calculated with GraphPad-PRISM 4.00 software. All determinations were performed in triplicate. Antiproliferative assays For the antiproliferative assay, we used sulforhodamine B (SRB), a colorimetric assay which estimates cell number by staining total cellular protein with the SRB dye, in order to assess cell growth inhibition [11]. The method used the same conditions as the cytotoxic assay except that medium was replaced with DMEM 10% SFB to induce cellular proliferation during extract treatments. After 48 h incubation, the medium was discarded and cells were fixed with 100 µl ice-cold 40% trichloroacetic acid (TCA, Aldrich Chemical). Thereafter, the cells were incubated at 4°C for 1 h and plates were washed five times with cold water. The excess water was drained off and the plates were left to dry, 50 µL of SRB stain (10 mg w/v in 1% acetic acid) were added to each well for 30 min. Finally, they were washed with 50 mL 1% acetic acid, rinsed four times until dye adhering to the cells was observed. The OD was measured at 540 nm using an ELISA reader (model 450, BIO-RAD, USA). Untreated cells were used as negative control. The IC50 value, i.e., the concentration of organic extract that produced a 50% reduction of the surviving fraction, was calculated using GraphPad-PRISM 4.00 software. MDCK cell line was used to evaluate the selectivity and cytotoxicity of the alga extracts. All determinations were performed in triplicate. Agarose gel electrophoresis Induction of apoptosis on cancer cells was assayed by DNA agarose gel electrophoresis. Cancer cells at exponential phase were exposed to 50 µg mL-1 of organic extract for 48 h. The cells were harvested, washed twice with PBS, and DNA was extracted using the DNA Ladder Detection Kit (Biovision). Ifosfamide (Sigma), an antitumoral drug, was used as a positive control while untreated SiHa cells were used as negative control. DNA was analyzed by electrophoresis at 75 mA on 8 g L-1 agarose gel containing 0.5 mg L-1 ethidium bromide. After 1.5 h, the gel was visualized and photographed under transmission UV light. Total phenolic content Total phenolic content of algal extracts was determined spectrophotometrically using Folin-Ciocalteu reagent [12]. First, 20 mg of the dry extract was diluted with methanol (3 mL). Aliquots of the diluted extracts (0.1 mL) were transferred into the test tubes; 2.9 mL of distilled water and 0.5 mL of Folin-Ciocalteu reagent were added. After 10 min, 1.5 mL of 20% sodium carbonate solution was added, mixed thoroughly and allowed to stand at room temperature in the dark for 1 h. Absorbance was measured at 725 nm, total phenolic content (expressed in % of dry weight) was calculated based on a standard curve of phloroglucinol. Total lipid content Total lipids were determined according to a previously reported method [13]. Alga extract (20 mg) was homogenized with a mixture of H2O, methanol and chloroform (1:1:9 v/v). The chloroform layer containing dissolved lipids was collected, dried with nitrogen and saponified with 1.2M NaOH. Fatty acids were converted to methyl esters with 0.6 mL 10M HCl and 1 mL of 12% boron trichloride in methanol at 80°C for 60 min. After methylation 1 mL of hexane: diethylether (1:1) and 3 mL of 0.3M NaOH were added, dried with nitrogen and recovered with hexane. The total content of fatty acid methyl esters was analyzed by gas chromatography (Hewlett Packard 6890 Plus with Supelco SP2560™ biscianopropyl polysiloxane capillary column 100 m x 0.25 mm x 0.25 µm internal diameter). The column temperature programming was set from 140 (5 min) to 240 ºC (20 min) at a rate of 4°C/min. Injector and detector temperature was 260ºC. Helium was used as the carrier gas at a flow rate of 1.1 mL min-1. Fatty acid methyl esters were identified by comparing their retention times with those of standard samples. The lipid analyses were carried out in duplicate and the results expressed as percentage of alga extract dry weight (% dry wt). Statistical analysis Data were analyzed with a software (GraphPad Software Inc, San Diego, CA). The dose– response curves (variable slope) were fitted with the algorithm: Y= Emin + {(Emax_Emin) / (1 + 10(Log ED50 -Log D) Hill slope)}. Statistical analysis was performed with parametric tests, because variances were homogeneous between groups. Unpaired Student’s t-test (two-tailed) was applied when only two groups were compared. One-way ANOVA followed by post hoc Tukey’s test was used to assess the differences when three or more groups were simultaneously compared. Values in text and figures are expressed as mean ± SD. RESULTS The antiproliferative activities of algal extracts increased when the wild alga was cultured under experimental conditions. The organic extract of P. dumetosus collected in the wild (time 0) showed antiproliferative activity on SiHa, HeLa, Hep-2 and KB cells with IC50 of 360.1 ± 1.3, 224.6 ± 3.2, 76.0± 2.2 and 44.8 ± 1.6 µg mL-1 respectively (Table 1). In general, all P. dumetosus extracts obtained after cultivation treatments exhibited an increase in biological activity, as they showed increased inhibitory effects with a reduction in the initial IC50, thus suggesting that culture conditions enhanced extract biological activity. This effect was most evident after 10 days of cultivation, particularly on SiHa, Hep-2 and KB cells. The antiproliferative activity of P. dumetosus extracts on SiHa cancerous cell line was significantly lower than the initial growth inhibition activity (IC50 360.10 µg mL 1), with a reduction of 80 – 90 % of the initial IC50. In Hep-2 cells, the inhibition of cell growth was in the IC50 range of 11.7 -17.4 µg mL-1, representing a reduction of 77.0 -84.5 % from the original extract activity (IC50, 76 µg mL-1). For KB cell line the inhibition of cell growth was in the IC50 range of 19.3 -29.9 µg mL-1, representing a reduction of 33.3 -56.9 % from the control (IC50 44.8 µg mL-1). Moreover, activities against KB cells declined at 20 and 30 days. HeLa cell line was more susceptible to extracts obtained from P. dumetosus after cultivation at day 10 and 20, compared to the original extract. The inhibition of cell growth was in the IC50 range of 70.6 -91.2 µg mL-1 and 50.5 -68.8 µg mL-1 at 20 and 30 days, respectively. These values represent a reduction of 59.4 -68.6 and 69.4-77.5 %, respectively, compared to the control (IC50, 224.60 µg mL-1). Penicillus dumetosus extracts showed reduced cytotoxicity (CC50 , 300 ± 2.3 µg mL-1) including the alga extract before cultivation (Table 2). The extracts obtained from P. dumetosus cultured under different DNA ladder pattern of apoptosis induced by Penicillus dumetosus extract from treatment A after 10 days of culture (50 µg mL-1) for each cell line is shown in Figure 1. This pattern was similar to the positive control. Formation of DNA fragments of oligonucleosomal size (180-200 bp) is characteristic of apoptosis in cells. The extract obtained from wild collected P. dumetosus before cultivation (time 0) showed a total phenol content of 2.9 ± 0.17 % dry wt. This value was considered as 100 % to provide a basis of comparison with the extracts from the light-treated algae. No significant differences in phenolic content were found for the different applied irradiances nor in relation to culture time, i.e., 10, 20, 30 days (one-way ANOVA, F(11, 24) = 1.24, p = 0.08, Figure 2a). The extract obtained from freshly collected P. dumetosus before cultivation (time 0) showed a total lipid content of 57.7 ± 1.07 % This value was considered as 100 % in order to provide a basis of comparison with the extracts from the light-treated algae. Total lipid content of organic extracts varied in relation to time and light availability (Figure 2b). Lipid content in P. dumetosus subjected to different levels of irradiance, after 10 days, decreased to approximately 66 – 72 % in relation to control. After 20 days, P. dumetosus increased its lipid content, particularly under B treatment (140 %); however, after 30 days, lipid content decreased by 71 to 94 %. GC analysis revealed that saturated fatty acids (SAFA) were dominated (77.4 %) while monounsaturated fatty acid (MUFA) and polyunsaturated fatty acid (PUFA) constituted 12.3 and 10.3 %, respectively (Figure 2c). In general, after cultivation, SAFA content decreased from 10 to 54 % in relation to control while MUFA content remained unchanged throughout the test period and cultivation treatment. On the other hand, PUFA content increased 2 - 4 times in plants subjected to culture treatments, compared to the control. The fatty acid profile of the different extracts obtained in this study showed that the predominant SAFA was palmitic acid (16:0), while in the MUFA fraction, palmitoleic acid (PA; C16:1) and oleic acid (OA; C18:1) predominated. In general, P. dumetosus extracts had high levels of PUFA (omega-3 and omega-6) that increased after 10 days of cultivation, approximately four times in irradiance treatments A and B, and two times in irradiance treatments C and D. DISCUSSION Extracts obtained from P. dumetosus cultured at different light irradiances displayed varying antiproliferative activities against the cancer cell lines employed in this study. In other alga models, such as the cyanophyte Scytonema, increasing irradiance gradually increased antibiotic production [14]. Of all macroalgae, green alga are the least prolific producers of natural products, with less than 300 known compounds and only a handful of new secondary metabolites reported each year [1]. Tropical green alga in the order Bryopsidales, including the genus Penicillus are noted for the production of sesqui and diterpenoids that could be responsible of this antiproliferative activity [2]. Triterpene esters capisterones A and B derived from this green alga have also shown potent inhibition on marine fungi [4]. Recent studies have suggested that polyphenol content in alga extracts could be responsible for antiproliferative activity on cancer cell lines [8]. Moreover, phenolic compounds including phlorotannins can induce oxidative stress in cancer cells [15]. Previous studies have reported similar phenolic content of P. dumetosus and Penicillus pyriformis to be 2.20 ± 0.06 and 1.30 ± 0.16 % dry wt, respectively [16]. In this study the phenol content did not change over time or under the light treatment conditions tested and was not related to increased antiproliferative activity of the extracts. Fatty acids have also been identified as bioactive compounds in marine algae. Eicosapentanoic acid (EPA) from the marine diatom Phaeodactylum tricornutum has been proven to have antibacterial activity against diverse pathogenic bacteria [17]. Moreover, a variety of fatty acids and their derivatives have antiproliferative effects in different cancer cell models. In particular, polyunsaturated fatty acids (PUFAs) have been demonstrated to reduce proliferation and induce cell death in studies with leukemia cell lines [9]. In this study, we observed an increase in the PUFA content of P. dumetosus extracts during cultivation. In particular, EPA content increased four-fold after 30 days cultivation under darkness, whereas, other fatty acids reported to have biological activity, such as PA, did not change significantly during the cultivation period or as a result of light treatment. In cultured P. dumetosus, a combination of high nitrogen and reduced temperature observed during the 10-day period probably had an effect on increased PUFA content. According to the American National Cancer Institute, the IC50 limit to consider a crude extract promising for further purification is < 30 µg mL-1 [18]. Based on this criterion, after 10 days of cultivation, all P. dumetosus extracts increased their activity against Hep-2 and KB cells. Lower cytotoxicity in normal cells, compared to cancer cells, was also found in P. dumetosus extracts, a prerequisite for any potential extract for further isolation of bioactive compounds. On the other hand, apoptosis gives an idea about the effectiveness of anticancer therapy; some anticancer drugs show their anticancer activity by inducing apoptosis of cancer cells [19]. DNA fragments of oligonucleosomal size (180 -200 bp) are a characteristic of apoptosis in cells. The results of the present study strongly suggest that the suppressive effect of P. dumetosus extracts on cancer cell proliferation is related to apoptotic body formation. The exact underlying mechanisms of the antitumor activity of algae are as varied as the chemistry of the various secondary metabolites involved. In this regard, caulepenyne from Caulerpa taxifolia has been shown to inhibit proliferation of the fibroblastic cell line BHK21/C13 and SK-N-SH, and its cytotoxicity has also been demonstrated in various tumor cell lines [20]. Several sulphated algal polysaccharides may also reduce cell proliferation of cancerous colonic epithelial cells [21]. While the metabolites responsible for the antiproliferative activities in P. dumetosus have not been chemically characterized, the data indicate that multiple secondary compounds with biological activity may be produced at a given time. This has been proven in higher plants where the magnitude of pharmacological effects may vary depending on where the plant was grown, when it was harvested and how long it was stored [3]. CONCLUSION The results of this study indicate that the metabolism of some active compounds of P. dumetosus are substantially influenced by changes in light treatments, and hence have an influence on the production of metabolites that render them biologically active. Extracts obtained from P. dumetosus after 10 days of culture under conditions described in this study exhibited increased antiproliferative activity and induced apoptosis on SiHa, HeLa, Hep-2 and KB cells. Although we observed culture conditions and time influenced lipid content and composition, the compound responsible for the antiproliferative activity of P. dumetosus extracts remains to be identified. ACKNOWLEDGMENT This work was supported by SEP-CONACYT 53687. Cinvestav financed a post-doctoral fellowship for R Moo-Puc. The authors thank C Chávez Quintal and ML Zaldivar Romero for technical assistance during analysis. REFERENCES
Copyright © 2011 - Pharmacotherapy Group, Faculty of Pharmacy, University of Benin, Benin City, 300001 Nigeria. The following images related to this document are available:Photo images[pr11025t2.jpg] [pr11025f1.jpg] [pr11025t1.jpg] [pr11025f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}