 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
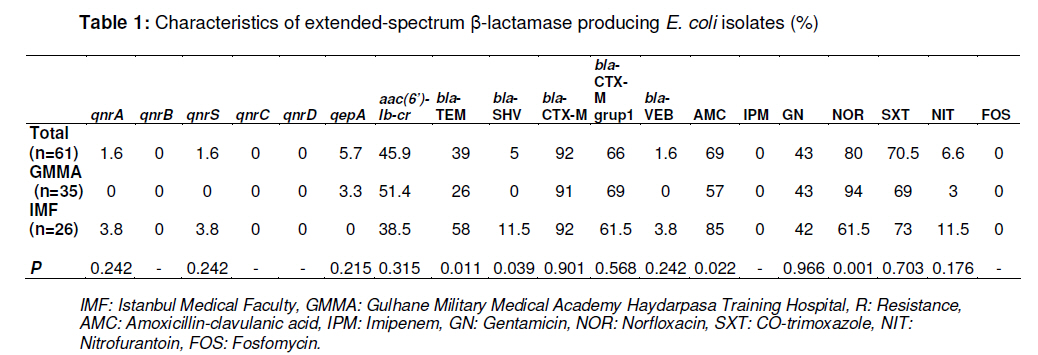
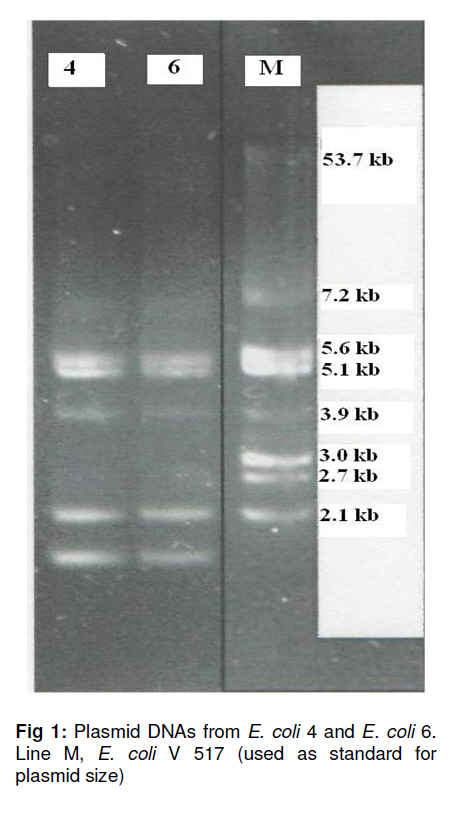
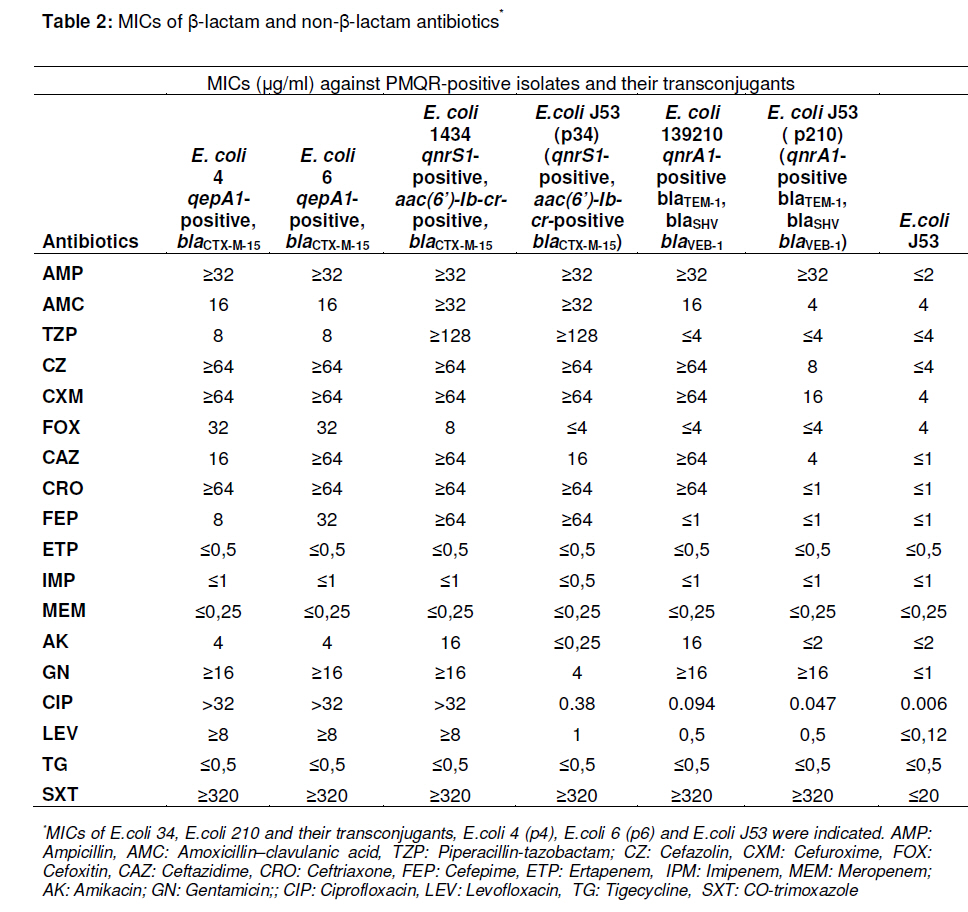
Tropical Journal of Pharmaceutical Research, Vol. 10, No. 3, June, 2011, pp. 325-333 Research Article Plasmid-Mediated Quinolone Resistance Genes in Escherichia coli Urinary Isolates from Two Teaching Hospitals in Turkey: Coexistence of TEM, SHV, CTX-M and VEB-1 Type β-lactamases Hasan Nazik1*, Bayhan Bektöre2, Betigül Öngen1, Mehmet İlktaç1, Mustafa Özyurt2, Nuray Kuvat1, Orhan Baylan2, Hatun Keküllüoğlu1, Tunçer Haznedaroğlu2 and Fatih Mehmet Keleşoğlu3 1Istanbul Medical Faculty, Department of Microbiology and Clinical Microbiology, Received: 5 October 2010 Revised accepted: 16 April 2011 Code Number: pr11042 DOI: 10.4314/tjpr.v10i3.9 Abstract Purpose: To evaluate the occurrence of plasmid-mediated quinolone resistance (PMQR) genes and the prevalence of extended spectrum β-lactamase (ESBL) types in Escherichia coli clinical isolates. Keywords: β-lactamase, Quinolone resistance, aac(6’)-Ib-cr, CTX-M-15, VEB-1 INTRODUCTION Although bacterial resistance to quinolones is usually due to chromosomally encoded mechanisms, it can also originate from plasmid-mediated genes. After the identification of qnr determinants, which protect target enzymes against quinolone inhibition, two other mechanisms have now been described: the qepA gene encodes an efflux pump, which confers reduced susceptibility on hydrophobic fluoroquinolones such as norfloxacin and ciprofloxacin; and the aac(6’)-Ib-cr gene, which encodes modified aminoglycosideacetylating enzymes and can inactivate both aminoglycosides and fluoroquinolones [1-3]. Over the last decade, plasmid-mediated quinolone resistance (PMQR), particularly among the various species of Enterobacteriacae, has been increasingly reported from many regions of the world. Plasmids carrying genes may contribute to the development of higher levels of fluoroquinolone resistance and may pose a threat by allowing the rapid spread of resistance among organisms. Although these PMQR genes have been associated with low levels of quinolone resistance, it could cause high-level quinolone resistance by facilitating the selection of chromosomal mutations. Several studies have demonstrated that most qnr-positive enterobacterial isolates are associated with extended spectrum βlactamases (ESBLs), including TEM, SHV, VEB, and CTX-M types, which are generally located on plasmids that are highly transferable and may harbor resistance genes to several different groups of antibiotics [4]. Today, many antibiotics, such as β-lactams and fluoroquinolones, which are widely prescribed by clinicians for the treatment of urinary tract E. coli infection, are in limited use. The production of ESBLs and PMQR proteins are a cause for concern. The ciprofloxacin resistance and ESBL rates are high in E. coli isolates from Turkey, being 40 -42% and 28.7 -32.1 %, respectively [5,6]. However, significantly lower ciprofloxacin resistance and ESBL rates among E. coli in North America, Latin America, and Europe were observed -4.5 -1.9 %, 7.1 -9.0 %, and 5.0 - 5.4 %, respectively [7]. Data on the prevalence of ESBL, especially CTX-M and VEB types, are limited in Turkey. Although the prevalence of PMQR genes, qnrA, qnrB, qnrS, aac(6’)-Ib-cr, and recently qepA and associated ESBLs have been reported, the prevalence of qnrC and qnrD is unknown [811]. The aim of this study was to determine the prevalence of PMQR genes and ESBL types in clinical urinary isolates of E. coli collected from two large teaching hospitals located in the European and Asian parts of Istanbul in Turkey. EXPERIMENTAL Bacterial isolates A total of 61 consecutive non-repetitive ESBL-producing E. coli strains, from the years 2008 and 2009, were collected from the Microbiology Laboratories of two teaching hospitals in Istanbul, Turkey. One of the hospitals, Istanbul Medical Faculty (IMF, 1.750 beds), is located in the European part of Istanbul, while the other, Gulhane Military Medical Academy Haydarpasa Training Hospital (GMMA, 1000 beds), is in the Asian section of Istanbul. Isolates from Istanbul Medical Faculty (n = 26) and from Gulhane Military Medical Academy Haydarpasa Training Hospital (n = 35) were included. The isolates were collected from urine specimens and isolated and identified with the aid of Chromogenic medium and Vitek 2 System (bioMérieux, France). Antimicrobial susceptibility and synergy testing Individual strains were tested based on the recommendations of the Clinical and Laboratory Standards Institute (CLSI), using the Kirby–Bauer disc diffusion method for susceptibility [12]. The double disc synergy test with cefotaxime and ceftazidime was used for screening ESBL production. The following antibiotic discs (Oxoid, Hampshire, UK) were purchased and used, as instructed by the manufacturer: amoxicillin-clavulanic acid (20/10 µg), imipenem (10 µg), gentamicin (10 µg), norfloxacin (10 µg), co-trimoxazole (1.25/23.75 µg), nitrofurantoin (300 µg), and fosfomycin (200 µg). E. coli 25922 was used as control strain. For PMQR-positive isolates and their transconjugants, the minimal inhibitory concentration (MIC) of ampicillin, amoxicillin-clavulanic acid, piperacillinclavulanic acid, cefazolin, cefuroxime, cefoxitin, ceftazidime, ceftriaxone, cefepime, ertapenem, imipenem, meropenem, amikacin, gentamicin, levofloxacin, tigecycline, and co-trimoxazole were determined by Vitek 2 System. The MICs of ciprofloxacin were determined by E-test method (AB, Biodisk, Solna, Sweden). Enterobacterial repetitive consensus PCR (ERIC-PCR) The Enterobacterial Repetitive Intergenic Concensus (ERIC)-PCR with ERIC1 and ERIC2 primers was used to analyze the epidemiological relationship between PMQRpositive E. coli isolates. Cycling conditions were as follows: 5 min at 94 °C; 40 cycles of 1 min at 94 °C, 1 min at 36 °C, 2 min at 72 °C; and final extension of 10 min at 72 °C. The PCR products were separated by electrophoresis in 1.5 % agarose gel and visualized on a UV transilluminator, and fingerprints were compared [13]. Transferability of PMQR genes and plasmid analysis Conjugation experiments with an azideresistant E. coli J53 (AzR) as the recipient were performed in liquid culture media, as described previously [14]. Transconjugants were selected on trypticase soy agar plates containing sodium azide (100 µg/ml) for counter selection and amoxicillin (100 µg/ml), cefotaxime (8 µg/ml), ceftazidime (8 µg/ml), nalidixic acid (16 µg/ml). The High Pure Plasmid Isolation Plasmid DNA Kit (Roche, Mannheim, Germany) was used for the extraction of plasmid DNA. E. coli V517 cells harboring plasmids of 54.4, 7.1, 5.6, 5.2, 3.0, 2.7, and 2.1 kb were used as the size marker for the plasmids. The presence of transferred PMQR genes and related ESBLs were confirmed by PCR. Characterization of ESBL and PMQR genes and sequencing DNA extraction was performed, as described previously. Briefly, bacterial colonies were suspended in 2 ml centrifuge tubes and then centrifuged at 12,000 g. The pellets were washed in 750 µl TE buffer (10 mM Tris HCl, pH 8.0, 1 mM EDTA) and then boiled for 10 min in 500 µl TE buffer and centrifuged. The supernatants were stored at -20 °C prior to subsequent DNA amplification [9]. The blaTEM, blaSHV, blaCTX-M, and blaVEB genes were investigated by PCR, as previously described [15,16]. A multiplex PCR was performed to detect qnrA, qnrB, and qnrS, as previously described by Cattoir et al [17]. PCR amplification of qnrC, qnrD, qepA, and aac (6’)-Ib was carried out with specific primers and conditions [2,3,18]. The DNA for control for each specific gene region was included with each group of tested strains. After PCR amplifications, the products of aac(6’)-Ib positives were further analysed by digestion with BtsCI for detection of the –cr variant (New England Biolabs, Ipswich, MA, USA). The amplification products of PMQR and related β-lactamases were sequenced with an Applied Biosystems sequencer (ABI PRISM 310 Genetic Analyzer; Applied Biosystems, Foster City, CA, USA). The nucleotide and amino acid sequences were analyzed and compared by BLAST search (www.ncbi.nlm.nih.gov). Statistical analysis Statistical analysis was performed using SPSS for Windows, version 11.5 (SPSS, Inc, Chicago, IL, USA). Rates of resistance were compared using the Chi-square test. A pvalue of < 0.05 was considered to be statistically significant. RESULTS Antibiotic susceptibilities and the prevalence of PMQR determinants/βlactamase genes Table 1 shows the prevalence and susceptibility data. The strains displayed highest resistance to norfloxacin (80 %). The most active antibiotics were imipenem (100 %), fosfomycin (100 %) and nitrofurantoin (93.4 %). The rate of resistance to amoxicillin-clavulanic acid for the isolates of IMF was higher than that of the isolates of GMMA (p= 0.022); the opposite was found for the resistance to norfloxacin (p= 0.001). Co-resistance was identified in 96 % of the strains. The highest co-resistance was determined for norfloxacin and the most common two co-resistance phenotypes were amoxicillin-clavulanic acid/norfloxacin/cotrimoxazole (24.5 %) and amoxicillinclavulanic acid/gentamicin/norfloxacin/cotrimoxazole (15 %). The prevalence of PMQR genes for aac(6’)Ib-cr, qepA, qnrA, and qnrS were 45.9, 5.7, 1.6, and 1.6%, respectively. qnrA1 and qepA1 were detected alone in strains, but qnrS1 was co-expressed with aac(6’)-Ib-cr. All PMQR-positive isolates were resistant to norfloxacin, except the qnrA1-positive strain. In addition, norfloxacin resistance in aac(6’)Ib-cr-positive isolates (all were resistant) was significantly higher than in the aac(6’)-Ib-crnegative ones (p = 0.001). No isolates carrying the qnrB, qnrC, or qnrD genes were detected in this study (Table 1). The most prevalent ESBL type was CTX-M (92 %) (mostly CTX-M group 1 (66 %)), followed by TEM and SHV. Only one isolate harbored the VEB-1 type β-lactamase. Ciprofloxacinresistant E. coli 4 and E. coli 6 were isolated from patients with nephrolithiasis, who were operated in the same division of GMMA during the same period, and these were qepA1-positive. E. coli 4 was isolated 36 days after the operation and the patient was treated with fosfomycin. E. coli 6 was isolated from a patient who was admitted with high fever 11 days after the operation and treated with a imipenem-gentamicin combination. Ciprofloxacin-resistant E. coli34 (from a kidney transplant patient treated with piperacillin-tazobactam) and ciprofloxacinsusceptible E. coli 210 (from a patient who was born with premature rupture of membranes treated with an ampicillingentamicin combination) were isolated in different divisions of ITF. E. coli 24 harbored both qnrS1 and aac(6’)-Ib-cr, while qnrA1 was detected in E. coli 210. RAPD-PCR typing E. coli 4 and E. coli 6 have similar antibiotic patterns; i.e., they are resistant to amoxicillinclavulanic acid, cefriaxone, amikacin, gentamicin, norfloxacin, levofloxacin, and cotrimoxazole. RAPD-PCR typing was carried out on the four PMQR-positive isolates. The results showed that the qepA1-positive isolates were clonally related (data not shown). Characteristics of PMQR-positive isolates, transconjugants, and plasmid analysis PCR assays were used to detect βlactamase, and identified the blaCTX-M-15 gene in E. coli 4, E. coli 6, and E. coli 34, while blaTEM-1, blaSHV, and blaVEB-1 genes were detected in E. coli 210. Despite three separate attempts, conjugative assays failed with the E. coli 4 and E. coli 6 isolates. However, plasmid analysis demonstrated that both strains harbored multiple plasmids changing 1-7 kb (Figure 1). The pattern of susceptibility to the β-lactams of the transconjugants corresponded to the expression of clavulanic acid-inhibited ESBLs. Transfer of the qnrS1+aac(6’)-Ib-cr and blaCTX-M-15 of E. coli 34 and qnrA1, blaTEM1 , blaSHV , and blaVEB-1 gene of E. coli 210 to the azide-resistant E. coli J53 occurred during the conjugation experiments. Tranconjugants of E. coli 34 were resistant to amoxicillin-clavulanic acid, cefriaxone, amikacin, gentamicin, levofloxacin, and cotrimoxazole. Transconjugants of E. coli 210 were resistant to cefriaxone, gentamicin, and co-trimoxazole. The MICs of ciprofloxacin and other antibiotics for E. coli 34 and E. coli 210 parenteral isolates and their transconjugants are presented in Table 2. The MICs for ciprofloxacin were increased 63 and 8 times in the transconjugants of E. coli 34 and E. coli 210, respectively. DISCUSSION It is more difficult to treat ESBL-producing E. coli because most β-lactams are no longer therapeutic options. In particular, CTX-M type enzymes have emerged worldwide and have rapidly increased in E. coli isolated from both community and nosocomial settings [19]. The associated co-resistance of ESBL producers to different groups of antimicrobials, such as quinolones, sulfonamides, and aminoglycosides, is another issue of concern. Limited data have been reported on the epidemiology of E. coli that produce CTX-M type enzymes in Turkey. Two recent reports from Turkey have shown that the CTX-M enzyme is common among ESBL positive isolates (86.8 %) at our hospital (IMF) in Istanbul [20] and from patients with urinary tract infections (76.5%) [21]. The latter finding of the predominance of CTX-M type enzymes according to TEM and SHV types is reflected in our study, as well. These reports suggest that CTX-M type enzymes are more prevalent than other ESBLs in Turkey. Consistently, a high prevalence of CTX-M type enzymes has been reported in several studies from other countries. In addition, the present study demonstrates that the prevalence of VEB type β-lactamase is low (1.6 %). Over the past 10 years, PMQR has emerged as an important issue. Different rates of PMQR have been reported depending on the country of origin of the isolates [22]. E. coli carrying qnrA, qnrB, qnrS, and aac(6’)-Ib-cr genes have been previously reported in Turkey [8-10,23]. In the present study, the most prevalent PMQR determinant was aac(6’)-Ib-cr (45.9%). A low prevalence was detected for the other PMQR genes (0 -5.7 %). Many studies have demonstrated an association between TEM-SHV-CTX-M type β-lactamases and PMQR in Enterobacteriaceae [4]. In Turkey, as in previous reports, PMQR genes were mostly associated with TEM, SHV, and CTX-M type β-lactamases, such as the qnrS1+aac(6’)-Ib-cr -blaCTX-M-15 positive E. coli 34 strain in this study. The association of qnrA with VEB-1 type βlactamases was first investigated in a single Enterobacter cloacae isolate from France and in 11 out of 23 blaVEB-1 positive enterobacterial isolates from Thailand, by Poirel et al [16]. In addition, a qnrA-positive-Citrobacter freundii isolate that produces blaVEB-1 and blaOXA-48 has been reported in Turkey [8]. Here, in addition to the TEM and SHV type, a VEB type β-lactamase was detected, but distinct from that of a qnrApositive E. coli isolate from the same hospital. This finding showed that VEB-1 type βlactamase persists in microorganisms in Turkey. The present study also demonstrated the co-expression of the PMQR genes, similar to previous reports from France, UK, China, and Turkey [11,24-26]. The strain, qepA, was first identified in 2007 in two E. coli clinical isolates from Japan and Belgium [27], while a new variant (qepA2) has already been detected in France [28]. However, recently, a qepA producing E. coli strain possessing qnrB2 and aac(6’)-Ib-cr gene has been reported in Turkey [11]. In the present study, in addition to qnrA1, qnrS1 and aac(6’)-Ib-cr, qepA1-positive E. coli isolates that produce CTX-M-15 type βlactamase were indentified. No qepA was found in the isolates screened in ITF, in contrast to other hospitals located in the Asian part of Istanbul. Although these PMQR mechanisms are rare, the association of qepA1 with multi-drug resistant CTX-M-15 producing E. coli can be a cause for concern. There may be a rapid spread of E. coli, especially in hospital settings where various antimicrobials are largely used and thus may support the dissemination of these microorganisms. CONCLUSION This study documents the high prevalence aac(6’)-Ib-cr and CTX-M type enzymes in Turkey. In addition to aac(6’)-Ib-cr and CTX-M type enzymes, the qepA1 and VEB-1 type enzymes are alarming for Turkey. Our study confirms that CTX-M producing E. coli isolates from urine specimens are highly resistant/co-resistant to norfloxacin, cotrimoxazole, amoxicillin-clavulanic acid, and gentamicin. Empiric therapy with these antibiotics may not be adequately effective. However, fosfomycin and nitrofurantoin resistance rates seem low and they may be alternatives for therapy. The emergence of the combination of PMQR and ESBL compromise the usage of valuable antibiotics worldwide. Antibiotic resistance is a public health problem, which requires continuous surveillance, monitoring, and revision of the policy of antibiotic use. ACKNOWLEDGEMENT The authors thank Professor Patrice Nordmann and Laurent Poirel for kindly providing the qepA positive strain. We thank Professor Minggui Wang for kindly providing qnrC and qnrD. We are grateful to Lina Cavaco for her kind cooperation and agreement in supplying qnrD. REFERENCES
Copyright © 2011 - Pharmacotherapy Group, Faculty of Pharmacy, University of Benin, Benin City, 300001 Nigeria The following images related to this document are available:Photo images[pr11042t2.jpg] [pr11042t1.jpg] [pr11042f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}