 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
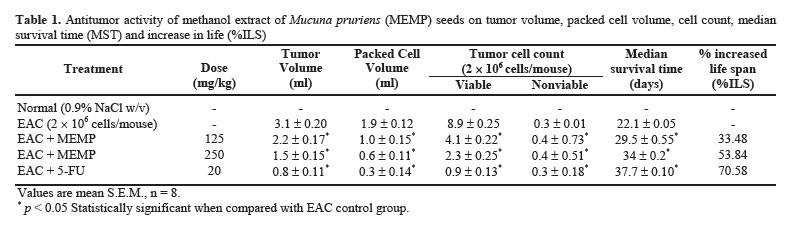
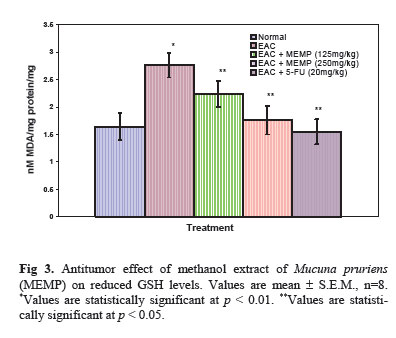
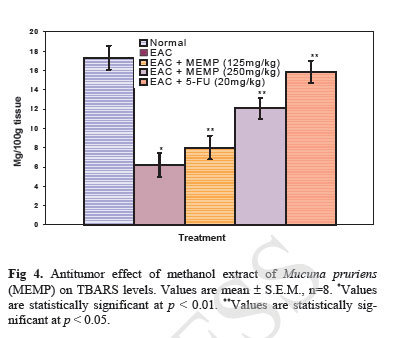
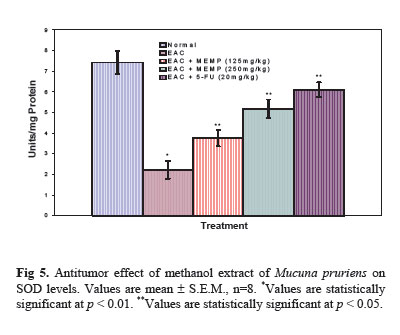
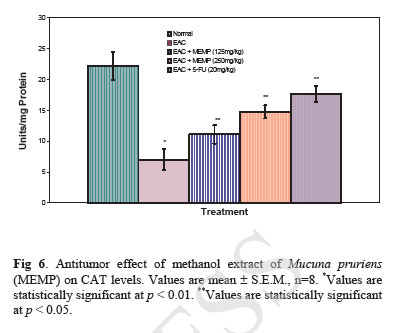
Iranian Journal of Pharmacology & Therapeutics, Vol. 4, No. 1, 2005, pp. 46-53 Antitumor Activity and in vivo Antioxidant Status of Mucuna pruriens (Fabaceae) Seeds against Ehrlich Ascites Carcinoma in Swiss Albino Mice YERRA RAJESHWAR, MALAYA GUPTA and UPAL KANTI MAZUMDER Division of Pharmaceutical Chemistry and Pharmacology, Department of Pharmaceutical Technology, Jadavpur Uni-versity, Kolkata, India. Address correspondence to: Prof. Upal Kanti Mazumder, Department of Pharmaceutical Technology, Jadavpur Univer-sity, Kolkata, India. E-mail: yrajeshwar@yahoo.co.in Received May 1, 2005; Code Number: pt05012 ABSTRACT The aim of the present study is to evaluate the antitumor effect and antioxidant role of Mucuna pruriens (Family: Fabaceae) against EAC bearing Swiss albino mice. The effect of methanol extract of Mucuna pruriens (MEMP) on tumor growth and host’s survival time was studied by the following parameters: tu-mor volume, packed cell volume, viable and non-viable cell count and life span of the host. MEMP was administered at a 125 and 250mg/kg b.w. once a day for 14 days, after 24 h of tumor inoculation. De-crease in tumor volume, packed cell volume, and viable cell count were observed in MEMP treated ani-mals when compared to EAC treated animals. Treatment with MEMP at a dose of 125 and 250mg/kg in-creased the mean survival time to 29.5 ± 0.55 and 34 ± 0.2 days respectively. The extract also decreased the body weight of the EAC tumor bearing mice. Hematological studies reveal that the Hb content was decreased in EAC treated mouse, whereas restoration to near normal levels was observed in extract treated animals. There was a significant decrease in RBC count and increase in WBC counts in extract treated animals when compared to EAC treated animals. The study was also extended to estimate the liver biochemical parameters such as LPO, GSH, and antioxidant enzymes like SOD, CAT etc. Treatment with MEMP decreased the levels of lipid peroxidation and increased the levels of glutathione (GSH), su-peroxide dismutase (SOD) and catalase (CAT). The results suggest that the methanol extract of Mucuna pruriens seeds exhibits significant antitumor and antioxidant effects in EAC bearing mice. Keywords: Mucuna pruriens seeds, Ehrlich ascites carcinoma, Anticancer activity, in vivo antioxidant activity Oxygen free radicals are formed in tissue cells by many endogenous and exogenous causes such as me-tabolism, chemicals, and ionizing radiation [ 1 ]. Oxygen free radicals may attack lipids and DNA giving rise to a large number of damaged products [ 2 ]. Iron is known to be involved in the generation of reactive oxygen species (ROS) and in the formation of highly toxic hydroxyl radicals from other active oxygen species such as hy-drogen peroxide [2-4]. The enhanced generation of ROS in vivo could be quite deleterious, since they are in-volved in mutagenesis, apoptosis, ageing, and carcino-genesis [ 4 ]. Free radicals also cause DNA strand breaks and chromosome deletions and rearrangements. Further, activated oxygen species most likely play an important role in tumor promotion and progression [ 5 ]. For these reasons, the search for antioxidants as cancer chemo-preventive agents is a continued process. Various epi-demiological, experimental, and metabolic studies have show that nutrition plays an important causative role in the initiation, promotion, and progression stages of sev-eral types of human cancers [ 6 , 7 ]. In addition to sub-stances that pose cancer risk, the human diet also con-tains vegetables, fruits, and beverages, which not only provide essential vitamins and minerals, but include important chemopreventive agents capable of protecting against some forms of human cancer [ 6 - 8 ]. Many cancer chemopreventive agents possess antioxidant potential [ 8 ]. Such chemopreventive agents are known as anticar-cinogens, and ideally they should be non-toxic. Mucuna pruriens (Fabaceae) is an established herbal drug in the Ayurvedic system of medicine used for the management of male infertility, nervous disorders and also as an aphrodisiac [ 9 ]. Its English name is cowhage and in Hindi it is known as Kinanchh or Kewach. It is an annual herbaceous twinning plant, found all over India, especially in the tropics [ 10 ]. Eighty percent of the total alcohol extract of the seeds shows the presence of 5-indolic compounds, especially tryptamine and 5-hydroxytryptamine (5-HT) [ 11 ]. Mucunine, mucu-nadine, prurienine and prurieninine are the four alka-loids, isolated from its seed extracts [ 12 ]. It is also rich in fatty content [ 13 ]. The powder of the seeds is clini-cally used for the management of hyperprolactinaemia and Parkinson’s disease, as it contains a high concentra-tion of L-DOPA [ 14 ]. Its seeds produce a hypoglycae-mic effect and the fruits showed a weak neuromuscular blocking effect in normal rats but not in alloxan treated rats [ 15 ]. It is known that ageing, nervous disorders, a decrease in performance and some other sexual disor-ders are mediated through free radicals [ 16 ]. Thus, there is a great possibility that this plant may act through the mechanism of free radical removal, in the management of the above disorders. Tripathi et al. used iron-induced lipid peroxidation in rat liver homogenate to investigate the chemical interaction of various phytochemicals with different species of free radicals in vitro [ 17 ]. Previously, we reported the antiepileptic and anti-neoplastic activity of methanol extract of Mucuna pru-riens root [ 18 ]. Our recent findings revealed that the methanol extract of Mucuna pruriens seeds showed significant in vitro antioxidant activity [ 19 ]. So far no reports are available on in vivo antioxidant status of this plant in EAC tumor bearing mice. Hence, we evaluated the in vivo antioxidant status of the methanol extract of Mucuna pruriens (MEMP) seeds in EAC tumor bearing mice. MATERIALS AND METHODS Chemicals Nitroblue tetrazolium (NBT), Thiobarbituric acid (TBA), Phenazonium metho sulphate and Nicotinamide adenine dinucleotide (NADH) were purchased from Loba Chemie, Bombay, India. 5,51-dithio bis 2-nitro benzoic acid (DTNB), Folin-Ciocalteau phenol and re-duced Glutathione were purchased from SISCO Re-search Laboratory, Bombay, India, and bovine serum albumin from Sigma Chemical Co., St. Louis, MO, USA. All other reagents used were of analytical grade. Preparation of Plant Material and Extraction The seeds of the plant Mucuna pruriens were pur-chased from the United Chemicals and Allied Products, Kolkata, West Bengal, India. The seeds of the plant were identified and authenticated by the Botanical Sur-vey of India (BSI), Kolkata, India. A voucher specimen (No. GPS-1) has been preserved in our laboratory for future reference. The seeds were dried under shade and powdered (particle size ~0.25 mm) using a laboratory mill. The seed powder of 2000 g was extracted with 1500 mL of petroleum benzine (60-80°C) followed by methanol at 24°C for 72 h by hot continuous extraction method. The solvent was evaporated under reduced pressure at 50°C and dried in vacuum (Yield: 15.75%, methanol extract). The dried extract thus obtained was dissolved in isotonic normal saline solution and used directly for the assessment of antitumor and antioxidant activities. Ethical Clearance Protocol used in this study for the use of mice as an animal model for cancer was approved by the Univer-sity Animal Ethical Committee. Experimental Animals Male Swiss albino mice of about 8 weeks of age with an average body weight of 24 ± 2 g were used for the experiment. The animals were bred and brought up in our laboratory facility with 12-h cycles of light and dark at 23°C. They were fed standard laboratory diet and were given sterilized water ad libitum. Tumor Cells Ehrlich ascites carcinoma (EAC) cells were obtained from Chittaranjan National Cancer Institute, Kolkata, India. The EAC cells were maintained in vivo in Swiss albino mice, by intraperitoneal (i.p.) transplantation of 2 × 106 cells/mouse after every 10 days. EAC cells 9 days old were used for the screening of the MEMP. Experimental Protocol Male Swiss albino mice were divided in to five groups of eight animals (n=8) each. The MEMP was dissolved in isotonic saline (0.9% NaCl w/v.) solution and used directly in the assay. EAC cells were collected from the donor mouse and were suspended in sterile isotonic saline. The viable EAC cells were counted (Trypan blue indicator) under the microscope and were adjusted at 2 × 106 cells/mL. 0.1 mL of EAC cells per 10g body weight of the animals was injected (i.p.) on day zero (day 0). A day of incubation was allowed for multiplication of the cells. Fourteen doses of the MEMP (125 mg and 250 mg/kg, 0.1 mL/10g body weight) and 5-Fluorouracil (20 mg/kg body weight) were injected i.p. from the first day up to the 14th day with 24-h inter-vals. Control animals received only vehicle (isotonic saline solution). Food and water were withheld 18 h before sacrificing the animals. On day 15, half of the animals (n = 4) in each cage were killed and the remain-ing animals were kept to observe the life span of the hosts. 5-Fluorouracil (5-FU) at a dose level of 20 mg/kg body weight was used as standard. The anti-tumor activity of the methanol extract of Mucuna pruriens (MEMP) was measured in EAC ani-mals with respect to the following parameters:
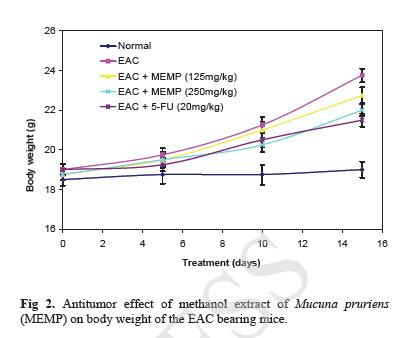
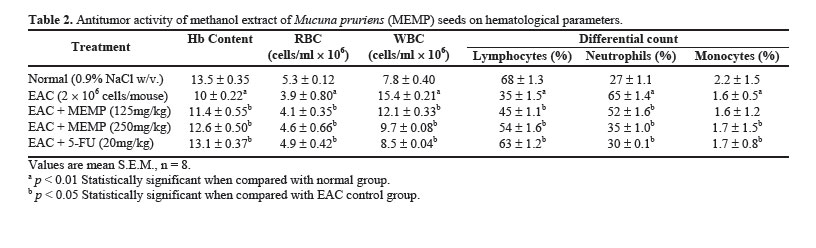
Acute Toxicity Study Albino male mice of 10 animals per group and weighing 20-25 g were administered graded doses (250-4000 mg/kg body weight, i.p.) of the methanol extract of Mucuna pruriens (MEMP). After administration of the MEMP the mice were observed for toxic effects after 48 h treatment. The toxicological effects were ob-served in terms of mortality and expressed as LD50. The number of animals dying during the period was noted [ 20 ]. The LD50 of the extract was calculated by the method of Litchfield and Wilcoxon [ 21 ]. Hematological Parameters At the end of the experimental period, all mice were killed the next day after an over night fast by decapita-tion. Blood was collected from freely flowing tail vein and used for the estimation Hemoglobin (Hb) content, red blood cell count (RBC) [ 22 ] and white blood cell count (WBC) [ 23 ]. WBC differential count was carried out from Leishman stained blood smears [ 24 ]. Estimation of in vivo Antioxidants After collecting the blood samples, the mice were killed by cervical dislocation. The liver was excised, rinsed in ice-cold normal saline solution followed by cold 0.15 M Tris-HCl (pH 7.4), blotted dried and weighed. A 10% w/v homogenate was prepared in 0.15 M Tris-HCl buffer and was used for the estimation of lipid peroxidation (LPO) and reduced glutathione (GSH). The rest of the homogenate was centrifuged at 1500 rpm for 15 min at 4°C. The supernatant thus ob-tained was used for the estimation of superoxide dismu-tase (SOD), catalase (CAT) and total protein. Estimation of Lipid Peroxidation (LPO) The levels of Thiobarbituric acid reactive substances (TBARS) in the liver was measured by the method of Ohkawa et al. (1979) [ 25 ] as a marker for lipid peroxi-dation. A mixture of 0.4 mL of 10% liver homogenate, 1.5 mL of 8.1% sodium dodecyl sulphate (SDS), 1.5 mL of 20% acetate buffer (pH 3.5) and 1.5 mL of 0.8% TBA solution was heated at 95°C for 1 h. After cooling, 5.0 mL of n-butanol-pyridine (15:1) was added, and the absorbance of the n-butanol-pyridine layer was meas-ured at 532 nm. Estimation of Reduced GSH The tissue GSH was determined by the method of Beutler and Kelly [ 26 ]. Virtually all the non-protein sulfhydryl groups of tissues are in the form of reduced GSH. 0.2 mL of tissue homogenate was mixed with 1.8 mL of EDTA solution. To this 3.0 mL precipitating reagent (after precipitating proteins with TCA) was added, mixed thoroughly and kept for 5 min before cen-trifugation. To 2.0 mL of the filtrate, 4.0 mL of 0.3-M disodium hydrogen phosphate solution and 1.0 mL of DTNB (5, 5-dithio bis 2-nitro benzoic acid) reagent were added and the absorbance read at 412nm. Assay of SOD The activity of SOD in tissue was assayed by the method of Kakkar [ 27]. The assay mixture contained 1.2 ml sodium pyrophosphate buffer (pH 8.3, 0.025 mol/L), 0.1ml phenazine methosulphate (186 mmol/L), 0.3 ml nitroblue tetrazolium (300 mmol/L), 0.2ml NADH (780 mmol/L) and diluted enzyme preparation and water in a total volume of 3 ml. After incubation at 30°C for 90 sec, the reaction was terminated by the addition of 1.0ml of glacial acetic acid. The reaction mixture was stirred vigorously and shaken with 4.0 ml n-butanol. The color intensity of the chromogen in the butanol layer was measured at 560 nm against n-butanol. Assay of CAT Catalase was assayed according to the method of Maehly and Chance [ 28]. The estimation was done spectrophotometrically following the decrease in absorbance at 230 nm. The tissue was homogenized in M/150 phosphate buffer (pH 7.0) at 4°C and centrifuged at 5000 rpm. The reaction mixture contained 0.01 M phosphate buffer (pH 7.0), 2 mM H2O2 and the enzyme extract. The specific activity of catalase is expressed in terms of units/mg protein. A unit is defined as the velocity constant per second. Estimation of Total Proteins The protein content of tissue homogenates was measured by the method of Lowry [ 29]. 0.5 ml of tissue homogenate was mixed with 0.5 ml of 10% TCA and centrifuged for 10 min. The precipitate obtained was dissolved in 1.0 ml of 0.1 N NaOH. From this an aliquot was taken for protein estimation. 0.1 ml of aliquot was mixed with 5.0 ml of alkaline copper reagent and allowed to stand at room temperature for 10 min. 0.5 ml of Folin’s phenol reagent was added and the blue color developed was read after 20 min at 640 nm. Statistical Analysis The experimental results were expressed as mean ± S.E.M. Data was assessed by ANOVA followed by the Students t-test, p-value < 0.05 was considered as statistically significant. RESULTS Acute Toxicity LD50 value of the methanol extract of Mucuna pruriens (MEMP) was evaluated in Swiss albino mice. The drug was administered intraperitoneally. As shown in the Fig 1, the LD50 value of the MEMP was found to be 1333.52 mg/kg body weight. Tumor growth response of MEMP on tumor and packed cell volume, viable and non-viable cell count and % increase in life span Antitumor activity of MEMP against EAC tumor bearing mice was assessed by the parameters such as tumor volume, packed cell volume, cell count (viable and non-viable), mean survival time and % increase of life span. The results are shown in Table 1. The tumor volume, packed cell volume and viable cell count were found to be significantly (p < 0.01) increased and nonviable cell count was significantly (p < 0.01) low in EAC control animals when compared with normal control animals. Administration of MEMP at the dose of 125 and 250 mg/kg significantly (p < 0.05) decreased the tumor volume, packed cell volume and viable cell count. Non-viable cell count was significantly (p < 0.05) higher in MEMP treated animals when compared with EAC control animals. Furthermore, the median survival time was increased to 29.5 ± 0.55 (%ILS = 33.48) and 34 ± 0.2 (%ILS = 53.84) on administration of MEMP at 125 and 250 mg/kg respectively. Finally, the change in body weights of the animals suggests the tumor growth inhibiting property of MEMP. All these results clearly indicate that the MEMP has a remarkable capacity to inhibit the growth of solid tumor induced by EAC cell line in a dose-dependent manner in experimental animals. (Fig 2) Hematological Parameters Hematological parameters (Table 2) of tumor bearing mice on day 14 were found to be significantly altered compared to the normal group. The total WBC count was found to be increased with a reduction of Hb content of RBC. The total number of RBC showed a modest change. In differential count of WBC, the percent of neutrophils increased while the lymphocyte count decreased. At the same time interval on MEMP 250 mg/kg treatment restored all the altered hematological parameters to almost near normal. MEMP 125 mg/kg treatment also recovered these altered depleted parameters towards normal though MEMP 250 mg treatment was found to be more effective. Antitumor Effect of MEMP on TBARS Levels ROS formed in cancer tissues results in lipid peroxidation and subsequently to increase in malondialdehyde (MDA) level. Fig 3 depicts the levels of TBARS in liver tissue of experimental animals. In the present study, the levels of MDA were significantly (p < 0.01) increased in EAC control animals when compared with normal control animals. After treatment with MEMP at 125 mg and 250 mg/kg and 5-FU at 20 mg/kg significantly (p< 0.05) reduced the MDA levels when compared with EAC control animals. Results were expressed as nmoles MDA/mg proteins/ml. Antitumor Effect of MEMP on Reduced GSH Levels Fig 4 illustrates the levels of reduced GSH in experimental groups. The levels of reduced GSH were significantly (p < 0.01) decreased in EAC control group when compared with normal control group. The levels of reduced GSH were found to be increased on administration of MEMP at 125 mg and 250 mg/kg and 5-FU at 20 mg/kg when compared with EAC control group. Antitumor Effect of MEMP on SOD Levels Fig 5 shows the activity of SOD in liver tissue of experimental groups. There was a significant (p < 0.01) reduction in the levels of liver SOD in EAC control animals. Administration of MEMP at 125 mg and 250 mg/kg and 5-FU 20 mg/kg increased the levels significantly (p < 0.05) as compared with EAC control animals. Antitumor Effect of MEMP on CAT Levels Fig 6 demonstrates the activity of catalase (CAT) in liver tissue of experimental mice. A significant (p< 0.01) reduction was observed in the activity of catalase in EAC control groups. Feeding with MEMP at 125 mg and 250 mg/kg and 5-FU 20 mg/kg increased the levels significantly (p < 0.05) as compared with EAC control mice. DISCUSSION The present investigation was carried out to evaluate the antitumor activity and antioxidant status of methanol extract of Mucuna pruriens (MEMP) in EAC tumor bearing mice. The MEMP treated animals at the doses of 125 and 250 mg/kg significantly inhibited the tumor volume, packed cell volume, tumor (viable) cell count, and brought back the hematological parameters to more or less normal levels. The extract also restored the hepatic lipid peroxidation and free radical scavenging enzyme GSH as well as antioxidant enzymes such as SOD and CAT in tumor-bearing mice to near normal levels. In EAC tumor bearing mice, a regular rapid increase in ascitic tumor volume was observed. Ascitic fluid is the direct nutritional source for tumor cells and a rapid increase in ascitic fluid with tumor growth would be a means to meet the nutritional requirement of tumor cells [ 30]. Treatment with MEMP inhibited the tumor volume, viable tumor cell count, and increased the life span of the tumor bearing mice. The reliable criteria for judging the value of any anticancer drug are the prolongation of the life span of animals [ 31]. It may be concluded that MEMP by decreasing the nutritional fluid volume and arresting the tumor growth increases the life span of EAC-bearing mice. Thus, MEMP has antitumor activity against EAC bearing mice. Usually, in cancer chemotherapy the major problems that are being encountered are of myelosuppression and anemia [ 32, 33]. The anemia encountered in tumor bearing mice is mainly due to reduction in RBC or hemoglobin percentage, and this may occur either due to iron deficiency or due to hemolytic or myelopathic conditions [ 34]. Treatment with MEMP brought back the hemoglobin (Hb) content, RBC and WBC count more or less to normal levels. This clearly indicates that MEMP possess protective action on the hemopoietic system. Malondialdehyde (MDA) is formed during oxidative degeneration as a product of free oxygen radicals [ 35], which is accepted as an indicator of lipid peroxidation [ 36]. MDA, the end product of lipid peroxidation, was reported to be higher in cancer tissues than in nondiseased organ [ 37]. Our findings indicate that TBARS levels in the tested cancerous tissues are higher than those in normal tissues. These results are in agreement with the published data [ 38, 39]. This emphasizes the reduction in free radical yield and the subsequent decrease in harm and damage to the cell membrane and decrease in MDA production. Glutathione, a potent inhibitor of the neoplastic process, plays an important role in the endogenous antioxidant system. It is found in particularly high concentration in the liver and is known to have a key function in the protective process. Excessive production of free radicals resulted in oxidative stress, which leads to damage to macromolecules, for example, lipid peroxidation in vivo [ 40]. It was also reported that the presence of tumors in the human body or in experimental animals is known to affect many functions of the vital organs, especially the liver, even when the site of the tumor does not interfere directly with organ function [ 41]. In our study, GSH levels in experimental mice were found to be significantly lower than that in the EAC control mice. SOD, CAT, and glutathione peroxides are involved in the clearance of superoxide and hydrogen peroxide (H2O2 ). SOD catalyses the diminution of superoxide into H2O2 , which has to be eliminated by glutathione peroxidase and/or catalase [ 42]. Further, it has been reported that a decrease in SOD activity in EAC bearing mice may be due to loss of Mn2+ containing SOD activity in EAC cells and the loss of mitochondria, leading to a decrease in total SOD activity in the liver [ 43]. A small amount of catalase (CAT) in tumor cells was reported [ 43]. The inhibition of SOD and CAT activities as a result of tumor growth were also reported [ 44]. Similar findings were observed in our present study in EAC bearing mice. The administration of MEMP at two different doses significantly increased the SOD and CAT levels in a dose dependent manner. It was reported that plant-derived extracts containing antioxidant principles showed cytotoxicity towards tumor cells [ 45] and antitumor activity in experimental animals [ 46]. Antitumor activity of these antioxidants is either through induction of apoptosis [ 47] or by inhibition of neovascularization [ 48]. The implication of free radicals in tumors is well documented [ 49, 50]. The free radical hypothesis supported the fact that the antioxidants effectively inhibit the tumor, and the observed properties may be attributed to the antioxidant and antitumor principles present in the plant extract. In conclusion, the present study demonstrates that the methanol extract of Mucuna pruriens (MEMP) increased the life span of EAC tumor bearing mice and decreased lipid peroxidation and thereby augmented the endogenous antioxidant enzymes in the liver. All these parameters suggest that the methanol extract of Mucuna pruriens seeds exhibits potential antitumor and antioxidant activities. REFERENCES
Copyright © 2005 by Razi Institute for Drug Research (RIDR) |
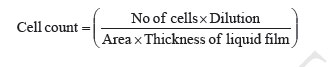

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}