 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Succinic Acid Monoethyl Ester and Metformin Regulates Carbohydrate Metabolic Enzymes and Improves Glycemic Control in Streptozotocin-Nicotinamide Induced Type 2 Diabetic Rats LEELAVINOTHAN PARI and RAMALINGAM SARAVANAN Department of Biochemistry and Biotechnology, Faculty of Science, Annamalai University, Annamalainagar, Tamil Nadu, India. Address correspondence to: Dr. L Pari, Department of Bio-chemistry and Biotechnology, Faculty of Science, Annamalai University, Annamalai Nagar-608 002, Tamil Nadu, India. Tel: + 914144 238343. Fax: + 914144 238145. E-mail: paribalaji@gmail.com Received September 9, 2005; Revised February 28, 2006; Accepted February 29, 2006 Code Number: pt05028 ABSTRACT Objective. Succnic acid mono ethyl ester (EMS) was recently proposed as an insulinotropic agent for the treatment of non-insulin dependent diabetes mellitus. In the present study the effect of EMS and Met-formin on the activities of carbohydrate metabolic enzymes in streptozotocin–nicotinamide induced type 2 diabeteic model was investigated. Keywords: Succinic Acid Monoethyl Ester, Nicotinamide, Metformin, Carbohydrate enzymes, Diabetes mellitus Type 2 diabetes is a chronic metabolic disorder characterized by abnormalities in carbohydrate and lipid metabolism [ 1 ]. It represents a heterogeneous group of disorders having hyperglycemia, which is due to im-paired carbohydrates (glucose) utilization resulting from a defective or deficient insulin secretory response. The liver plays a pivotal role in glucose and lipid homeosta-sis [ 2 ]. In experimental diabetes, enzymes of glucose metabolism are markedly altered and produce hypergly-cemia, which leads to pathogenesis of diabetic compli-cations [ 3 ] Glucose homeostasis involves the coordi-nated regulation of several metabolic pathways includ-ing gluconeogenesis and glycolysis. A number of therapeutic tools for the treatment of non-insulin dependent diabetes mellitus are currently available. A wide variety of structurally distinct mole-cules stimulate insulin secretion from pancreatic β cells by different mechanism of action. The ester carboxylic nutrient succinic acid (mono ethyl ester) derivative is a low toxicity drug (Fig 1) [ 5 ] and has attracted considerable interest for more than a decade as possible thera-peutic agent for the treatment of type 2 diabetes. This is based on the ability of EMS to act as potent stimulator of insulin secretion. It has previously been shown that succinic acid ester can be taken up and metabolized by pancreatic β cells, leading to increased pro-insulin bio-synthesis [ 6 ], insulin secretion and lower the blood glu-cose [ 7 , 8 ]. Its ester also protects pancreatic islets in vivo and in vitro against diabetogenic agents streptozotocin [ 9 ], interleukin 1β [ 10 ], and nitric oxide donor (NO) [ 11 ]. EMS is a suitable nutrient both to cover the energy need of hepatocytes and act as a gluconeogenic precursors [ 12 ]. Generally non-glucidic nutrients control hypergly-cemia, resulting in improved overall glycemic control in patients with type 2 diabetes. Metformin is an oral hypoglycemic agent, which be-longs to the class known as the biguanides. Chemically it is N-N dimethylimidodicarbonimidic diamide [ 13 ]. Metformin is now widely used as one of the mainstays in the management of type 2 diabetes. Metformin re-duces fasting plasma glucose concentration by reducing rate of hepatic glucose production via gluconeogenesis and glycogenolysis. Metformin improves glycemic con-trol as monotherapy and in combination with other oral antidiabetic agents, such as sulfonylureas and thiazolidine diones [ 14 ]. To our knowledge there are no other available re-ports on the effect of nonglucidic nutrient EMS on enzymes of hepatic glucose metabolism in streptozotocin-nicotinamide induced type 2 diabetes. Therefore the aim of the present study was to explore the effect of EMS on key hepatic enzyme in diabetic rats. The effect of EMS was compared with conventional antidiabetic agent met-formin. MATERIALS AND METHODS Drug and Chemicals Succinic acid monoethyl ester, and all other bio-chemicals and chemicals used in this experiment were purchased from Sigma chemical company Inc., St Louis, MO, USA. The chemicals were of analytical grade. Animals Healthy male albino Wistar strain rats (200-220g body weight) obtained from Central Animal House, Rajah Muthiah Medical College, Annamalai University were used in the present study. The rats were fed on pellet diet (Hindustan Lever Limited, Mumbai, India) and water ad libitum. The rats used in the present study were maintained in accordance with the guidelines of the National Institute of Nutrition, Indian Council of Medical Research, Hyderabad, India and the study ap-proved by the ethical committee (Vide. No:285,2005), Annamalai University. Experimental Induction of Type 2 Diabetes in Rats Non-Insulin dependent diabetes mellitus (NIDDM) was induced in overnight fasted rats by a single intrap-eritonial injection of 45 mg/kg streptozotocin, 15 min after the i.p administration of 110 mg/kg body weight of nicotinamide. Streptozotocin (STZ) was dissolved in citrate buffer (pH 4.5) and nicotinamide was dissolved in normal saline. Hyperglycemia was confirmed by the elevated glucose levels in plasma, determined at 72 h and then on day 7 after injection. The rats found with permanent NIDDM were used for the study [ 15 ]. Experimental Procedure In the experiment, a total of 42 rats (36 diabetic sur-viving rats, 6 control rats). Were used the rats were di-vided into seven groups of six rats each.
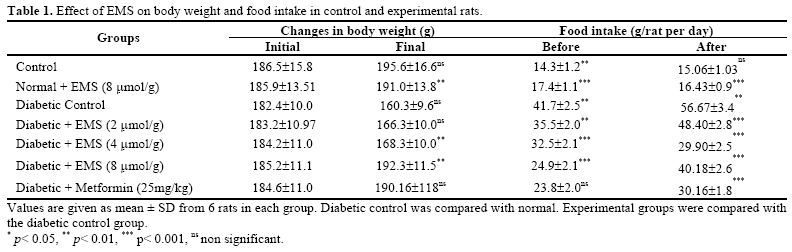
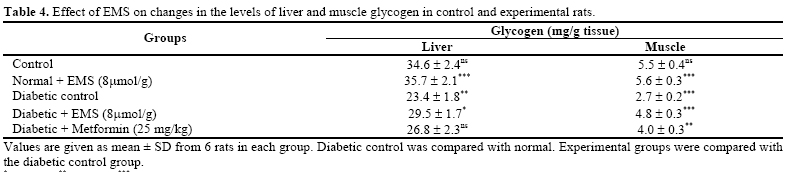
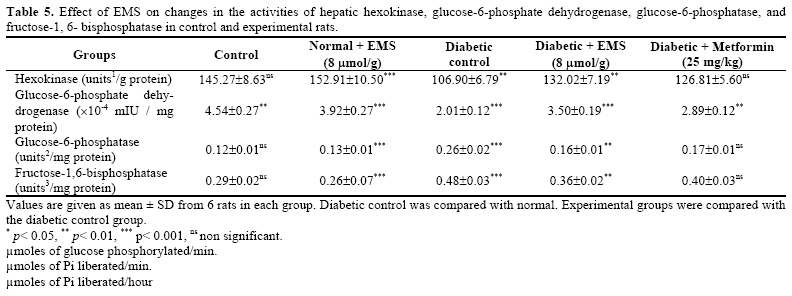
At the end of experimental period, the rats were de-prived of food overnight and blood was collected in a tube containing potassium oxalate and sodium fluoride for the estimation of blood glucose, hemoglobin and glycosylated hemoglobin. Plasma was separated for the assay of insulin. Liver was dissected out, washed in ice-cold saline, patted dry and weighed. Analytical Methods Determination of Plasma Glucose and Insulin. Plasma glucose was estimated calorimetrically using commercial diagnostic kits (Sigma Diagnostics (I) Pvt Ltd, Baroda, India) [ 17 ]. Plasma insulin was assayed using an enzyme linked immunosorbent assay (ELISA) kit (Boeheringer–Mannheim, Germany). Hemoglobin was estimated by using the cyanmethemoglobin method described by Drabkin and Austin [ 18 ]. Glycosylated hemoglobin was estimated according to the method of Sudhakar Nayak and Pattabiraman [ 19 ] with modifica-tions according to Bannon [ 20 ]. Determination of Liver Carbohydrate Enzymes. Glucose-6-phosphatase and fructose-1, 6-bisphosphatase were assayed according to the method of Koida and Oda [ 21 ] and Gancedo and Gancedo [ 22 ] respectively and the inorganic phosphorus (Pi) liberated was estimated by the method of Fiske and Subbarow [ 23 ]. Hexokinase and glucose-6-phosphate dehydro-genase were determined by the method of Brandstrup et al [ 24 ] and Ellis and Kirkman [ 25 ] respectively. Determination of Liver and Muscle Glycogen. Liver and muscle glycogen was estimated by the method of Morales et al. [ 26 ]. Statistical Analysis. All the grouped data were sta-tistically evaluated and the significance of various treatments was calculated using Student’s t-test. All the results were expressed as mean ± S.D. RESULTS Body Weight and Food Intake The changes in the body weight and food intake in control and experimental rats are represented in Table 1 . The body weights in EMS and metformin treated dia-betic rats increased significantly at the end of the ex-perimental period when compared with diabetic control rats. Food intake significantly increased in diabetic rats and it was significantly reduced in EMS and metformin treated groups compared to diabetic control rats. Plasma Glucose and Insulin Levels Table 2 demonstrates the level of plasma glucose and insulin in control and experimental animals. The level of plasma glucose was significantly increased whereas plasma insulin was significantly decreased in streptozotocin-nicotinamide diabetic rats. The administration of EMS significantly reversed the changes in a dose dependent manner. EMS at a dose of 8 μmol/g body weight showed a highly significant effect com-pared to 2 and 4μ mol/g body weight. Administration of EMS was compared with metformin, a references drug. The effect of EMS at a dose 8μmol/g body weights was used for further biochemical analysis. Hemoglobin and Glycosylated Hemoglobin Levels Table 3 shows the level of hemoglobin and glycosylated hemoglobin of different experimental groups. The diabetic rats showed a significant decrease in the level of total hemoglobin and a significant increase in the level of glycosylated hemoglobin. The administration of EMS and metformin to diabetic rats reversed the changes in total hemoglobin and glycosylated hemoglo-bin. Liver and Muscle Glycogen Levels Table 4 shows the changes in the level of liver and muscle glycogen of control and experimental rats. There was a significant reduction in liver and muscle glycogen of STZ-nicotinamide diabetic rats. Administration of EMS and metformin significantly increased the level of liver and muscle glycogen. Hepatic Carbohydrate Enzymes Levels Table 5 shows the changes in the activities of he-patic gluconeogenic enzymes (glucose-6-phosphatase and fructose 1, 6-bisphosphatase), hexokinase, and glu-cose-6-phosaphate dehydrogenase in different experi-mental groups. The activities of hepatic gluconeogenic enzymes were significantly increased whereas hexokinase and glucose-6-phosaphate dehydrogenase were significantly decreased in STZ-nicotinamide dia-betic rats. Administration of EMS and metformin to diabetic rats reversed the changes in the activities of these hepatic enzymes to almost control levels. DISCUSSION The ester of selected carboxylic metabolites, which are mediators in the Krebs cycle or their precursors such as pyruvic acid, succinic acid, and glutamic acid are currently under investigation as potent insulinotropic tools in the treatment of non insulin dependent diabetes [ 27 ]. The insulinotropic capacity of succinic acid monoethyl ester was first disclosed in pancreatic islets [ 28 ] .The nutritional value of this ester when infused to starved rats has been recently documented [ 29 ]. The fundamental mechanism underlying hyperglycemia in diabetes mellitus involves over-production (excessive hepatic glycogenolysis and gluconeogenesis) and de-creased utilization of glucose by the tissues [ 30 ]. The administration of EMS and metformin to de-crease the increased blood glucose concentration to normal glycemic concentration is an essential trigger for the liver to revert its normal homeostasis during ex-perimental diabetes. Glucose is metabolized in the pan-creatic β cell via glycolysis in the cytosol resulting in pyruvate production. Pyruvate is then transported into the mitochondria for Krebs cycle metabolism supporting subsequent oxidative phosphorylation and ATP produc-tion [ 31 - 33 ]. Generation of ATP by increased glucose metabolism promotes a rise in the cytosolic ATP/ADP ratio, closure of the ATP dependent K+ channel, open-ing of the voltage sensisitivity Ca2+ channels and a sub-sequent increase in the cytosolic free calcium concentra-tion resulting in triggering of proinsulin synthesis and insulin release [ 6 , 7 ] promoted by EMS. Metformin re-duces fasting plasma glucose level by reducing rates of hepatic glucose production, [ 34 , 35 ] its effect on the relative contributions of hepatic glycogenolysis [ 36 , 37 ] and gluconeogenesis [ 38 ]. EMS treatment significantly decreases plasma glucose and increases plasma insulin level compared with metformin. EMS at 8μ mol/g body weight was significantly better than 2 and 4 μmol/g body weight; therefore high dose was used further biochemical studies. In uncontrolled or poorly controlled diabetes, there is an increased glycation of a number of proteins includ-ing hemoglobin and alpha-crystalline of lens [ 39 ]. Glycated hemoglobin has been found to be significantly increased in diabetic animals and this increase was directly proportional to fasting blood glucose [ 40 ]. Ane-mia is a much more common disease in type 2 diabetic patients, potentially contributing to the pathogenesis of diabetic complications. Hemoglobin was also recorded as a binary outcome for estimating the risks for anemia [ 41 ]. In the present study, the decreased level of hemo-globin indicates the anemia in STZ-nicotinamide dia-betic rats. Since during diabetes, the excess glucose transport in the blood reacts with hemoglobin to form glycosylated hemoglobin. Administration of EMS with metformin controls the glycation of hemoglobin by its normoglycemic activity and thus decreases the level of glycated hemoglobin in STZ-nicotinamide diabetic rats. In our study, hepatic and skeletal muscle glycogen content was reduced significantly in diabetic controls. Insulin is the main regulator of glycogenesis in muscle and liver. The decrease in hepatic and skeletal muscle glycogen contents in diabetic rats have been observed earlier by Pari and Latha [ 42 ]. The decrease in both muscle and hepatic glycogen observed in this study may be due to lack of insulin in the diabetic state and this type of results is probably due to the inactivation of gly-cogen synthetase system. EMS for 30 days in diabetic rats resulted in a significant elevation of liver and mus-cle glycogen levels. This shows the one possible way of antidiabetogenic action of this nonglucidic nutrient may be improvement of glycogenesis process in muscle and liver. Hexokinase and glucose-6-phosphate dehydrogenase activities have been observed to decrease in STZ dia-betic rats, which may be due to loss of insulin receptors [ 43 ], production of glycated proteins and formation of amadori product [ 44 ]. Hexokinase plays an important role in the maintenance of glucose homeostasis and all the cells that metabolize glucose by ATP to produce glucose-6-phosphate. Administration of EMS and met-formin to STZ diabetic rats resulted in a significant re-versal in the activity of hexokinase. The activity of glucose-6-phosphate dehydrogenase the first regularery enzyme of pentose phosphate path-way was found to be decreased in diabetic animals [ 45 ] and increased in EMS and Metformin treated animals. The activity was higher in comparison to untreated dia-betic animals indicating improvement in glucose utiliza-tion by this pathway. This may be attributed to the insu-lin secretory effect of EMS as glucose-6-phosphate de-hydrogenase has been reported to increase the supply of NADPH. The hepatic gluconeogenic enzymes, glucose-6-phosphatase and fructose-1, 6-bisphosphatase were increased significantly in diabetic rats [ 46 ] The increased activities of two gluconeogenic enzymes from liver may be due to the activation or increased synthesis of the enzymes contributing to the increased glucose produc-tion during diabetes, by liver, and EMS and metformin treatment may be primarily modulating and regulating the activities of the two gluconeogenic enzymes, either through regulation by cyclic AMP and any other metabolic activation or inhibition of glycolysis and glu-coneogenesis [ 47 ]. Administration of EMS and Metformin significantly decreased the activities of gluconeogenic enzymes in diabetic rats. The level of plasma insulin was found to increase significantly in diabetic rats treated with EMS, which may be a consequence for the significant reduc-tion in the level of gluconeogenic enzymes. The reduc-tion in the activities of gluconeogenic enzymes can re-sult in the decreased concentration of glucose in blood. In conclusion our result indicate that non-glucidic nutrient EMS possess antidiabetic action. The present investigation draws out a sequential metabolic correlation between increased glycolysis and decreased gluconeogenesis, increased hydrogen shuttle reaction and normal glycemia stimulated by succinic acid mono ethyl ester, which may have been the biochemical mechanism through which glucose homeostasis is regulated. ABBREVIATIONS EMS - Succinic acid monoethyl ester NIDDM - Non insulin dependent diabetes mellitus STZ - Streptozotocin bw - Body weight REFERENCES
Copyright © 2005 by Razi Institute for Drug Research (RIDR) The following images related to this document are available:Photo images[pt05028t4.jpg] [pt05028t3.jpg] [pt05028t1.jpg] [pt05028f1.jpg] [pt05028t5.jpg] [pt05028t2.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}