 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Electronic journal, URL - http://bioline.bdt.org.br/py
Tradeoff analysis in planning networks of protected areas for biodiversity conservation Gordon Brent Ingram Environmental Planning, 1230 Hamilton Street, 204 Vancouver, Canada V6B 2S8; e-mail gbingram@axionet.com
Code Number: PY97003
Size of Files:
Text: 113.3K
Graphics: Line Drawings (gif) - 31.0K
Key words: biological diversity conservation tradeoff analysis, networks of protected areas SUMMARY Much of modern nature conservation has been preoccupied with variations on the largely nineteenth century colonial notion of the national park. In this current period of tremendous rates of loss of habitat and biological resources, the pool of interventions available to the land use planner and manager must be expanded with underlying relationships and possibilities further explored. This paper outlines some of the tenets of the emerging theory of environmental planning for the conservation of biological diversity and considers the myriad of interventions available. Such measures are grouped into:
2. reserve management; and 3. regulation of land use external to protected areas.
2. total area of protected habitat and relatively "natural" or intact condition of those ecosystems within reserve boundaries; and 3. management within protected areas and regulation of land use outside of reserves.
INTRODUCTION As has been said in a thousand other articles over the last three decades, these are times of multiple crises of increasing loss of natural ecosystems and intensifying fragmentation, accelerating extirpations and extinctions of species, and mass loss of intra-specific variation. The purpose of this paper is to argue that the "paradigm" of "island biogeography" ^1 as the central notion in conservation biology must be re-examined and modified. I argue that for there to be adequate conservation of biological diversity we must move quickly from protected area planning, design and management frameworks, based on metaphors of discrete islands, to approaches recognizing archipelagos with dangerously fluctuating changing sea levels ^2. I also argue that there are usually far more options for effective conservation planning than are usually recognized in land management approaches based on notions of discreet habitat islands. Development of multiple scenarios for the identification of options for conservation and development, with various social, political, economic, and aesthetic processes, becomes as central to the long-term success of conservation initiatives as to the basic biological calculations for "minimum viable populations" (Shafer 1987). Ecosystem management across landscapes (Grumbine 1994) has emerged as the central concept, indeed something of a paradigm, for the conservation of such vague concepts as "biological diversity", often referred to as "biodiversity" (Roberts 1988, WRI/IUCN/UNEP 1992)^3. However, a theoretical framework to integrate conservation biology into environmental planning has been slow to emerge, in large part because the notion of biodiversity has been appropriated more by intellectual property law than as a basis for understanding the conservation of natural species, ecosystems, and processes (Athanasious 1996, p. 206). This paper explores some emergent concepts for conservation of biological diversity that re- examine and partially diverge from that of the island biogeography theory that emerged several decades ago^4. It explores the opportunities of the last decade, which has seen the emergence of the field of landscape ecology (Forman and Godron 1986) and its emphasis on relationships across varying spatial and temporal scales within broader regional and district "contexts." As has often been the case in the history of science, the development of new methodologies and techniques has preceded the means to define and verify desired conditions (Kuhn 1970, Feyerabend 1988). In most contemporary conservation planning and management, the goals and purposes for preservation of populations, ecological processes, and landscape attributes have not been clearly defined. In this vacuum, the fields of environmental planning and land management have absorbed and come to reflect operational concepts that roughly satisfy imperatives for such better conservation, particularly in terms of integration with economic development. But this common wisdom is still not supported by a verified body of knowledge that could, in turn, lay the basis for a theory of environmental planning for the conservation of biological diversity. A set of predicted "tradeoffs" ^5 with minimum levels of conservation for the ongoing persistence of local biological diversity has already been recognized without being fully described, confirmed, or debated. There surely must be more than one way to conserve biological diversity and respective "genetically coded functions" (Vogel & Ingram 1993) for particular areas. With accelerating ecosystem degradation, the extinction crisis, and genetic erosion, this notion poses a number of questions which are more than rhetorical. Three of these terms, "way," "conserve," and "biological diversity" embody conceptions with widely varied interpretations. In the mode of scientific discourse, so typical of our times, unresolved interpretations lead to a softening of the science. Consequently, regional conservation "simulation" ^6 has too often been relegated to technical aspects of "planning and management" even though the underlying assumptions of algorithms and models are heavily imbued with biases and short-term policy priorities.
Conservation biology, as a new science, has provided a home for many unresolved interdisciplinary questions but today still harbours reductionist biases - contrary to the arguments of some of its founders^7. As with much of current thought, under the rubric of "postmodernism," ^8 frameworks for environmental problem-solving are increasingly "site- specific" ^9 to the extent that such seemingly generic processes as "fragmentation" (Ingram 1992a) must be investigated in terms of the totality of the natural and social variables in particular areas. Population behaviour in either in situ baseline sites, or in more problematic ex situ laboratories, has been considered the most precise or reproducible data for development of models for conservation planning. But with networks of protected areas across landscapes, there are never two configurations that are alike and organisms can behave significantly differently under varying conditions. There is, consequently, a problem of extrapolation of biological data particularly as it is used to predict impacts related to the potential loss of local biological diversity. It is not so hard as we would like to believe. It is therefore often necessary to use more "soft," perhaps better thought as "flaccid," forms of data particularly in trying to grapple with cumulative change across landscapes with their inevitable social dimensions. This contradiction has emerged as possibilities and expectations for precision in conservation planning have increased, especially with the widespread use of geographic information systems and global positioning devices, and as the public and state will to intervene in ecosystems across landscapes has intensified. Recognition of imperatives for more comprehensive habitat conservation and biological diversity has stimulated new uses for environmental planning theory and methods. For example, Article 6 of the Convention on Biological Diversity (UNEP 1992), which came into force in December, 1993 ^10, includes a commitment to integrate concerns for biological diversity into land use planning and Article 7 talks of monitoring populations - presumably across landscapes. Article 8 embodies a commitment to establish and manage "systems" of protected areas. But even with such new instruments for cooperation and support of national agencies, crucial gaps in networks of protected areas will persist indefinitely in both the developing and the developed worlds. More recently, there have been increasingly critical discussions of the Convention and its emphasis which so far has been more on entrenchment of intellectual property rights than on actual habitat-based conservation (Athanasious 1996). The biodiversity concept, under the rubric of conservation biology, has emphasized a set of scientific principles and lines of inquiry, but there are still few widely recognized principles for practical linkages with regional planning. This design, planning and management aspect, the "art" of conservation biology, is still remarkably undeveloped. The weak basis for determining precise objectives for the management of configurations of "natural" sites across regions has, in itself, become an institutionalized obstacle to conservation of countless landscapes and biological resources. While today's accelerating rates of losses constitutes a crisis, it is crucial that we create the theoretical "space" to explore more systematic strategies, involving compilation of dependable and relevant information from increasingly diverse sources that is directly useful to decision-making. Such an expanded effort to develop theories of environmental planning for the conservation of local biological diversity has tremendous implications for evolving sampling frames, and inventorying and monitoring techniques. INVENTORYING POSSIBLE CONSERVATION INTERVENTIONS Identification of more quantitative, direct, and yet supple links between information and decision-making requires clearer assessments of the full range of interventions that are or could be available for habitat conservation in a particular locality. There have been only tentative efforts to catalogue and compare the planning measures which are available for particular regional administrative contexts, referred to by Barker (1982) as "fields of management." Little of this research has so far provided a basis for the construction of more comprehensive frameworks for decision-making both for conservation and expansion of land use. The ecosystem conservation strategies that have emerged around the various incarnations of the U.S. Endangered Species Act and the national strategies under the Convention on Biological Diversity come closest to providing a basis for the development of comprehensive and potentially preemptive strategies. These regional efforts have increasingly involved new sampling frames and spatial data compiled for multiple scales.
There have been two principal obstacles to comprehensive planning for biodiversity conservation. The first has been the lack of more regional and ecosystem-specific methods for identification of "indicators" of conservation of local biological diversity (Noss 1990, Ingram 1992b). The second has been the lack of a theory for relating the spatial and functional processes - an applied landscape ecology for biodiversity conservation (Ingram 1989; Diaz & Apostol 1992). The key information links between landscape ecology and conservation planning require spatial models with numerous linkages between natural and social conditions. The remainder of this article deals with this second problem by discerning some interrelationships of conservation interventions under the rubric of a more comprehensive paradigm of integrated resource management. Rarely can or should natural areas be protected solely because of requirement for maintenance of local biological diversity. There are usually additional concerns for resources and environmental costs, such as continued levels of production of certain species important for subsistence or as commodities, as well as non-market values related to recreation, heritage, and visual resources. There have been few theoretical discussions of the integration of an expanding array of noncommodity values into more comprehensive landscape planning (Ingram 1991). Even within ecological reserves and national parks, there is the issue of multiple use, such as balancing pressures for ecotourism and procurement of genetic resources. Particular conservation measures are never comprehensive in terms of system-wide or site-specific relationships and tend to favour the persistence of certain biological resources over others. This unevenness is the result of shifting and often erratic social priorities and acceptable levels of security in conservation.
Identification of relatively intrinsic relationships between potential combinations of conservation measures provides a better basis for organizing options for regimens of interventions into more politicized strategies. There must be more than one way to develop and maintain a system of protected areas that support minimum levels of conservation of biological diversity, even when particular objectives, standards, and social acceptable levels of risk vary radically with the extent of scientific information and priorities for social development. There should be, however, some underlying relationships, and respective categories of information and intervention that are more stable. Knowledge of the entire range of possible interventions, and respective relationships, is key to developing conservation programmes that are more comprehensive and adaptive (Hollings 1978). Three rough and overlapping categories of conservation interventions have emerged in the twentieth century:
2. protected area management; and 3. "regulation" ^11 of human activities outside of protected areas.
ALLOCATION OF AREAS FOR HABITAT PROTECTION Categories of protected areas vary greatly between jurisdictions. In some situations, particularly in the industrialized west, there are numerous possible designations. Each protected area embodies a unique set of purposes, agencies, and legislative frameworks. Each category of protected area has its own legislative, administrative and technical history and arena of response for countering aspects of habitat conversion or degradation. Certain types of protected areas have stronger and more defined administrative and legislative support than others.
There is an increasing awareness of the limitations of the national park ideal. In his post-colonial evaluation of national parks in Kenya, Lusigi (1978) provided the most powerful critique. Donna Haraway (1991) noted that,
"In establishing the game parks of Africa, European law turned indigenous human inhabitants of the `nature reserves' into poachers, invaders in their own terrain, or into part of the wildlife."
The rapid destruction of natural habitats, and the relatively slow rate of national park establishment, has prompted recognition that this category of protected area is rarely sufficient in terms of even selective preservation of alleles, genotypes, species, ecosystems, and mosaics of communities across entire regions. The "core", "buffer" (MAB 1974) and transition zones concepts were initial solutions to the need for prescriptive forms of conservation planning and management, particularly for biosphere reserves. In the last decade, more systematic, bioregional and site-based typologies of configurations of protected habitat across various natural and social edges and gradients have been identified (Schonewald-Cox et al. 1992; Buechner et al. 1992). A less orthodox approach to habitat allocation is the conservation easement (Barrett and Livermore 1983). Protection rests not in ownership per se, but rather in negotiated accords and collaborations between landowners and conservation groups. Subsequent stewardship responsibilities are provisional and often focus on control of human activities which threaten particular habitats and species. Joe Vogel (1994) has also proposed the concept of "genesteading" where private or more likely communal owners of natural ecosystems derive the resources to protect such areas through revenues from the transfer of genetic resources and intellectual property rights (The Crucible Group 1994). MANAGEMENT OF PROTECTED AREAS
With reserve management, we are confronted more fully with the dynamic nature of landscapes and ecosystems and by the vagueness of most goals for habitat conservation. In their evaluation of the effectiveness of conservation in national parks, Machliss and Tichnell (1985) spoke of the "historical lack of ecological management" which threatens the capacities of protected areas to maintain local biological diversity. However, the underlying lack of clear, comprehensive, and precise ecosystem management criteria threatens to be a more persistent problem.
Management consists of those conservation measures which are done after protected areas have been established within clearly delineated boundaries. Management is the modification of human-induced disturbance, substitution of natural processes, and as Usher (1973) noted, "is itself concerned with intervention within ecosystems, either to alter the status quo in a wanted direction, or to prevent the status quo from altering in an unwanted direction."
Control of human activities within protected areas is grounded in a long history of social conflict that continues to this day. There are dynamics between the cotters and the gentry, between local people and bureaucrats, between local and national governments, between provincial governments and federal governments, between rich and poor, between the "disempowered" and the powerful. Each decision over a protected area: its planning, design, and management, touch on these various, historically root conflicts and exists within broader political economies ^12. Park management has often involved policing and restraining activities, inevitably favouring some social groups and "stakeholders" (Ingram 1994b) over others. Management activities usually begin with the formulation of goals related to the changing conditions and distributions of species, habitats, features, sites, and landscape attributes. Management, therefore, is based on clarification of the operational and de facto "purposes" of particular reserves and the wide range of social needs for natural and exploited ecosystems and landscapes. Such specific purposes are ultimately based on economic, recreational and aesthetic values that are increasingly coming under the rubric of, but unfortunately often obfuscated by, broader concerns for conservation of biological diversity. MITIGATION OF IMPACTS OF ADJACENT LAND USE ON PROTECTED HABITAT "The success of a sanctuary depends largely upon what is done outside the sanctuary to make it a real and adequate haven for wildlife." Wright and Thompson 1935 ^13, as quoted by Shafer (1994). Mitigation for conservation of biological diversity can be defined as restrictions on human activities outside of protected areas in order to preclude negative impacts on populations and sites within reserves. In his essay on the range of legal tools for preservation of genetic diversity, De Klemm (1985) states that existing protected areas in themselves are not sufficient for effective nature conservation. There are a number of types of supplementary activities which can support habitat protection including general land use controls, permits, legal constraints, and administrative and economic incentives and disincentives. Mitigation measures involve the same dimensions, progressions of activities and levels of implementation as protected area allocation and management. However, the legal basis for mitigation, and thus its potential use and effectiveness, is often unresolved especially in terms of restrictions argued to be unfair "taking." This is especially the case where there are questions as to what constitutes the line between publicly and privately owned resources. Mitigation measures are underemployed in contemporary efforts for biological conservation. This is, in part, because the national park ideal has emphasized creation of supposedly secure and invulnerable islands of natural habitat even though, given the porosity of most landscapes, such discreetness rarely exists. A drawback to the allocation, management and mitigation trinity is that it highlights divisions rather than the fluid continuum of possible interventions for protection of habitat across landscapes. For example, it is not very important whether a conservation easement mechanism is considered a form of area allocation of protected habitat or the management of what might be a buffer in a larger network. What is significant is a specific measure's potential for relationships with other prescriptions across a landscape, over time. THE NOTION OF THE NETWORK IN HABITAT PROTECTION The metaphor of the discrete protected area as an island with sharp boundaries has given way to that of linked assemblages of concentric membranes. External boundaries function as filters rather than moats and are highly sensitive to the interplay of external and internal forces. For example, wolves can roam outside of reserve boundaries and air pollution intrudes inside. In their essay on the role of national parks in Italy, Giacomini and Romani (1978) described a shift from parks as "protectionist instruments" to "elements of total planning." They viewed parks as open systems and stated that, "attempts to create or operate on a park while considering it as a closed system are destined to failure."
In their multi-disciplinary boundary model, Schonewald-Cox and Bayless (1986) developed the concepts of the administrative boundary and the generated edge. The former is a mapped construct while the latter is often a volatile ecological gradient outside or inside of a reserve boundary. Too often disturbance-generated edges, "ecotones," extend well inside of administrative boundaries. Protected area management, particularly in the context of underdevelopment, is often focused on degraded edges that increasingly dominate habitat mosaics and threaten the viability of cores of protected habitat. Different combinations of allocation, management and mitigation measures can be ascertained to satisfy a set of objectives for maintenance of local biological diversity (Slayter 1974). Concentric membranes defining protected area edges often comprise "matrices" (Forman and Godron 1986, pp. 159 162 - 166) in mosaics of natural habitat in frontier landscapes. In more ideal situations it may be possible to confine impacts of land use activities to intensities, frequencies, and spatial distributions which are similar to those of less human-induced disturbance processes, thus limiting the dominance of "unnatural" edges. Research on ecotones by Hansen, di Castri and Naiman (1988) suggests that regional networks of protected habitat might be capable of functioning as "differentially-permeable membranes that facilitate some ecological flows but impede others."
Recognition of the supple interplay of possibilities for allocation, management, and mitigation in conservation planning and subsequent management has emerged over the last two decades. In their paper on conservation of macropods in reserves in western Australia, Main and Yadov (1971) first explored the relationship of reserve size to requirements for management. Almost a decade earlier, Leopold et al. (1963) provided the rationale for active management by suggesting that few of the national parks in the United States were sufficiently large to be self-regulatory ecological units. This early notion of tradeoffs in conservation planning was initially expanded by Pyle (1980) when he stated that, "The fewer and less intense the hostile pressure from outside the reserves, the larger the area set aside, the lower the number of visitors and the friendlier the social climate in which the reserve exists, the greater its defensibility and manageability."
TRADEOFF ANALYSIS FOR IDENTIFICATION OF OPTIONS FOR CONSERVATION INTERVENTIONS Rather than attempting to build networks of nature reserves as series of isolated ecological islands, it is more realistic to accept most boundaries as very porous and the total areas too small for long term viability. An alternative goal in conservation planning is to create an entire membrane that links natural "cores" through "buffers" (Dyer and Holland 1991) of less natural but regulated areas. There is always more than one way to conserve plant diversity and natural ecosystems and the differences between options involve various "tradeoffs" in land, management costs and security, with various implications for local social and economic development. This problem of integrating biodiversity conservation into social development was directly addressed in the Convention on Biological Diversity (Martinez, 1995), though since its ratification we still have only seen a modest degree of innovation and more effective problem-solving. One of the most difficult aspects of designing nature reserves is finding a set of boundaries and conservation commitments that can operate within the local political environment.
By recognizing as many possible tradeoffs as possible, there are better chances of finding a `scenario', a set of possibilities, that might work, or that might lead to the often poorly defined state of "sustainable development" and various forms of "ecological security" (Athanasiou 1996, pp. 8, 293 - 295). Assessment of these social and economic realities should be made early in the plant preserve design process as many species, no matter how rare or endangered, are often more adaptable and able to persist in a range of conservation regimens than adjacent human communities. Three sets of tradeoffs that are often most evident are:
2. between quality of the protected habitat and the extent of subsequent management and mitigation interventions in subsequent decades and centuries; and 3. between internal interventions, "management" and restrictions on land use external cores, often conceived of as "mitigation."
AN EXPANDED PARADIGM OF ENVIRONMENTAL PLANNING FOR THE CONSERVATION OF BIOLOGICAL DIVERSITY
All planning is based on forms of predictive models whether or not they are fully recognized or clearly quantified. A land use plan is the product of an uneven melange of objectives, criteria, and data. How these are worked to form certain strategies and agendas is the result of particular sets of projections and cause-effect linkages resulting from incomplete data. In comprehensive planning exercises, an array of quantitative and spatial models of specific relationships are linked and structured. But the structuring in planning for the conservation of biological diversity so far has been simplistic and rarely interdisciplinary. There is still an aversion to linking biological requirements, spatial relationships and social pressures. It is certainly possible to determine alternative sets of measures which could maintain minimum levels of local biological diversity. Such regimens organized into alternative scenarios can be based on levels of fitness, resilience and even intra-specific variation for specified periods. In order to produce spatial simulations, new kinds of environmental planning models are necessary. An underlying paradigm emerges based on tradeoffs between land allocation, habitat quality, and regulation of ecosystems, both inside and outside reserves, and centres on three postulates. We call this the tradeoffs in area / site quality / regulation paradigm (tasr). It can be characterized by an enhanced recognition of the myriad of possible configurations of habitat, regulation, and active ecosystem intervention that could maintain local biological diversity - each possibility with its own set of implications for social and economic development. Aspects of the tasr paradigm are already well- recognized in conservation planning and three of the major tenets have already emerged.
THE AREA / REGULATION POSTULATE
There are regional and district-wide relationships between the total area in protected habitat and the level and constitution of the social and ecosystem management that will be subsequently necessary to maintain the local biological diversity. This conception of "regulation" as conservation interventions not directly concerned with great allocation of area in protected habitat, involves both management within designated reserves and regulation of exterior land use around and between networks of protected areas. Greater regulation would allow for less total area in protected habitat in order to satisfy minimum conservation requirements for certain complexes of organisms and ecological processes. A greater spatial allocation of protected habitat could involve fewer requirements for regulation in order to accomplish the same conservation objectives. The basis for this postulate is that designation of protected area per se is not the only determinant as to whether respective habitat will continue to support prescribed sets of organisms over time. The viability of habitat, in terms of persistence of certain organisms, can diminish and even be permanently impaired without interventions which re-establish or substitute for natural processes and which minimize inevitable and often permanent human-induced change. But no amount of internal or external management can provide the basis for persistence of populations when certain population sizes, metapopulations, and landscape "processes" are not included within the spatial configurations of reserve networks. Janzen (1983) suggested such a minimum level of conservation when he noted that, "As areas of conserved pristine forest are reduced in size they are increasingly susceptible to significant immigration of animals and plants from nearby anthropogenic secondary successional habitats."
Such thresholds of susceptibility necessitate management and mitigation responses or greater total area in buffers of protected natural habitat. It is in consideration of the two end points, the extremes of this axis of minimum tradeoffs, that the relationships between total size, shape, configurations of area in protection, and regulatory interventions can be explored. The amount of flexibility available depends upon a range of natural and cultural relationships, and underlying cause-effect linkages, across landscapes. These relationships include population size, demographics, niche architecture, and habitat requirements for particular assemblages of species, the nature of episodic and stochastic change, and the resilience of particular ecosystems to specific disturbances. The area/regulation postulate promises some new opportunities for identifying more possibilities of protection of biological diversity compatible with priorities for social development. At one end of the axis of minimum tradeoffs, is the possibility of reserves so large that on-going requirements for protection of biological diversity through maintenance, regulation and constraints on adjacent land use, might be attractively low as averaged across landscapes. At the other extreme, land use expansions could be so well integrated into the buffer zones of networks of multi-purpose protected areas, with high levels of monitoring and constraints on human activities (Harmon 1994), that the total portion of the core areas could be modest. These relationships suggest that with tasr some ecosystems, landscapes and social pressures might inherently involve requirements for more total area in conservation than others. The social dimensions of the area/regulation postulate involve constraints on both traditional activities and modern land use, and involve various conservation stakeholders which can be local, national and international. The more negative and pervasive the prospective impacts of land use operations are, the greater are the requirements for regulatory intervention in land use or designation of more areas of protected habitat. There will always be some intrinsic biological, dynamic social and technical limits to the range of the possibilities of tradeoffs between allocation of reserves and regulation.
[b] - This point represents the total area of protected habitat necessary if there is minimal use of regulatory measures. [c] - This point represents the maximum amount of regulation and the minimum amount of protected habitat for the conservation of local biological diversity. Rather than wilderness, point [c] suggests highly manipulated habitat fragments. [d] - This point represents the total lack of both regulatory measures and protected habitat with accelerating losses in local biological diversity. [b - c axis] - This line represents the axis of the minimum tradeoffs between the least total area of a district necessary as protected habitat and the corresponding levels of regulation necessary for maintenance of local biological diversity. Below this threshold, loss of local biological diversity is inevitable. This line is probably curved with the nature of its shape, as are those of the other two axes of minimum tradeoffs described in the subsequent postulates, and are the results of regional and site-specific conditions resulting from a range of natural and social factors. [area between a, b, and c] - This area represents the subset of possible tradeoffs between protected area and regulation that can secure long-term conservation of local biological diversity. The points in this area represent viable regional conservation strategies in terms of both adequate total area in protection and regulatory measures. [area between d, b, and c] - This area represents configurations of habitat protection with inadequate levels of area and/or regulation, and represents ineffective strategies that could be associated with subsequent loss of elements of local biological diversity.
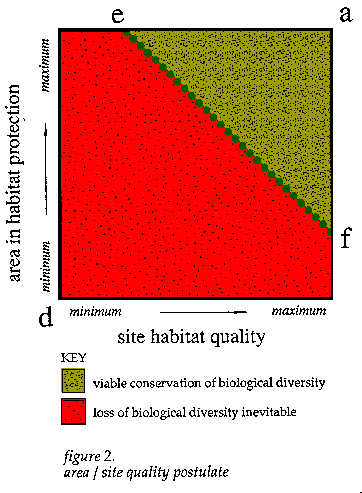
For conservation of local biological diversity, some forms of networks and configurations of sites could be more efficient in terms of allocation of space than others. Some sites have habitats which support lower densities of particular species. Some configurations involve more edge areas than others. This reality necessitates additional protected habitat for species which require habitat attributes associated with more mature successional phases. Larger areas are required to satisfy minimum requirements for populations across less productive, diverse or resilient mosaics. Some sites hold larger numbers of species and habitat attributes than others or contain elements that are particularly strategic, representative, rare or vulnerable. When sites of high concentrations are protected, it may be possible to keep the total area necessary in protected habitat lower than if conservation efforts had been focused on sites of relatively poorer quality in terms of supporting the persistence of local biological diversity. But there are fewer choices of sites for rare species and therefore tradeoffs are more limited. The area/site quality postulate lays the basis for identification of tradeoffs between the total area in protected habitat and the capabilities of sites to prescribed subsets of local organisms. If the natural areas with the highest quality of habitat are not chosen for protection, and instead are converted to consumptive uses, the remaining areas available for conservation will be generally less able to support the persistence of certain elements of local biological diversity. The total area necessary will need to be increased. This postulate has implications for the initial phases of conservation planning - the more key natural sites are destroyed before conservation planning, the lower the quality of habitat that will be available, and consequently the greater the possibility of a larger minimum area being necessary. The total area needed for a district network of protected areas would diminish if sites with configurations of more spatially efficient and secure areas of habitat were chosen early in land use planning. The extremes of this axis of the minimum area / site quality tradeoffs vary greatly depending on the requirements of targeted habitats and organisms, the distributions and aggregations of respective populations, and the nature of the pressures for expansion of destructive land uses. Determination of the axis of minimum tradeoffs between necessary area and site quality for particular biotic districts and social and economic contexts, is useful in a number of ways. The need for specific types and combinations of sites, in contrast to tracts which are chosen mainly out of their lack of consumptive economic values, can be highlighted. Potential sources of conflict, between pressures for extractive operations on certain sites and needs for biological conservation could be explored at phases sufficiently early in the planning process to allow for formulation of a great number of potentially viable alternatives, at least one of which is probably compatible with priorities for economic and social development.
Key: [a] - This point is the same as in figure 1, the area/ regulation postulate, with the maximum level of district area in habitat protection and with all key, high quality sites conserved. [d] - This point represents the lack of any area allocated to habitat protection with accelerating losses in local biological diversity. [e] - This point represents the largest portion of area necessary to allocate as protected habitat when configurations of only lower quality sites, in terms of spatial efficiency for persistence of local biological diversity, are available. [f] - This point represents the minimum total area of a district required for habitat protection when the highest quality sites, in terms of spatial efficiency for the persistence of local biological diversity, are chosen as protected areas. [e - f axis] - This line represents the set of tradeoffs, between the minimum portion of the area of a district in protected habitat and the corresponding requirements of the habitat quality of sites, in terms of supporting long-term persistence of local biological diversity. Below this threshold, loss of local biological diversity is inevitable. [area between a, e, and f] - This area represents the subset of tradeoffs between total area in habitat protection, quality of the sites, and the land use planning strategies associated with them, that provide the basis for viable conservation of local biological diversity. [area between d, e, and f] - The points here represent levels of both area in habitat protection and site quality that are insufficient for long-term persistence of local biological diversity.
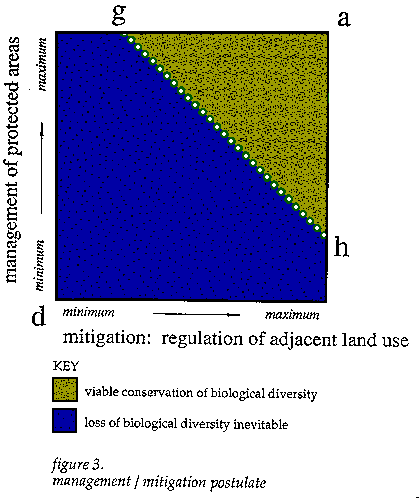
The third postulate results from an additional dimension of the first in terms of strategies for regulation. As part of a particular configuration of reserves and boundaries, there is a dynamic interrelationship between possibilities for regulation through management within protected areas and those for mitigation of the prospective negative impacts through regulation of land use outside of boundaries of protected areas. With more management inside of reserves, less mitigation would be necessary to regulate activities outside of reserves. The opposite could also be true. But there are limits to the plasticity of the tradeoffs between management and mitigation and active versus preclusive interventions, both in terms of intrinsic requirements for persistence of certain populations and what is socially, economically and fiscally feasible at a certain point in time. Management is implicitly an active affair which involves the substitution or reinforcement of natural processes or constraints on human activities. In contrast, mitigation involves specific forms of restraint in order to preclude certain negative aspects of expanding regional land use activities. While regulation of land use outside of reserves is by no means passive intervention, its implementation, at the administrative and legislative levels, tends to be preemptive. The temporal dimensions of management and mitigation measures can be quite different. The level of mitigation employed in the early phases of conservation programmes often determines the extent of management necessary for following years. However, the opposite is far less the case. Mitigation regimens largely determine the extent of the negative anthropogenic factors in the landscapes from which arise a large part of the necessity for management. In conservation planning, there is an opportunity to decide on the general balance of management and mitigation measures, or rather the preemptive versus the more adaptive and internalized strategies for maintenance of populations and habitat attributes. The extremes of this, the third dimension of the tradeoffs, are tied to the feasibility of various kinds of controls over sites, economic relationships, people and social dynamics in terms of imposition of limitations on land use activities. This relates back to the divergent implications of island versus membrane approaches for the design of regional networks of protected areas and subsequent management of land use expansion. The membrane approach works when land use throughout a district is carefully controlled or better self-managed to limit negative impacts.
Key: [a] - This is the same point described in the previous two postulates and in this case refers to the maximum level of conservation in terms of possible management and mitigation measures. [d] - This point represents the lack of management and mitigation measures, and the subsequent loss of biological diversity within protected areas. [g] - This point represents the maximum level of social and ecosystem management, in terms of forms and intensities, that can be employed within the protected habitat and the corresponding minimum requirements for regulation of land use activities in areas outside reserves. [h] - This point represents the maximum level of possible mitigation measures, for preclusion of certain human-induced disturbances outside of the boundaries of networks of reserves, and the corresponding minimum requirements for management within those reserves. [g - h axis] - This line represents the tradeoffs between minimum regimens of management and mitigation necessary to maintain biodiversity in a network of protected areas. Below this threshold, loss of local biological diversity is inevitable. [area between a, g, and h] - This area represents the potential tradeoffs between regimens of management and mitigation measures and associated land use that would support long-term conservation of local biological diversity. [area between d, g, and h] - The points here represent the various tradeoffs between management and mitigation measures, and their associated strategies, which would not lead to long-term persistence of local biological diversity.
How do we develop networks of protected areas in times of increasing globalization of capital and accelerating losses of biological diversity? The following are some "common sense" policy suggestions for any protected area planner and manager.
- No nature reserve is ever a true island. A nature reserve surrounded by farmland or city is not the same thing as a nature reserve surrounded by water. There are varying kinds of isolation that result from different kinds of land use. Particular levels of fragmentation reflect both intrinsic biological requirements and landscape processes on the one hand, and cumulative impacts from land use on the other. A reserve design embodies a strategy to control particular fragmentation and habitat degradation forces for a particular set of points and polygons in the landscape. - Islands never exist in isolation - all reserves must be designed as part of archipelagos. There is never much advantage in designing a single reserve. A group, a district-level archipelago, must be designed in conservation planning for local biodiversity. - All reserves are designed for certain conservation and economic development strategies (and good planning makes the respective goals transparent). There are always at least two alternative conservation/economic development strategies recognized in a landscape planning process. But instead of the preferred and the less-preferred strategies, at any point in time there should also be other possibilities that could be seriously explored, since they could become relevant with changing conditions.
The two persistent obstacles to a theory of environmental planning for the conservation of biological diversity are the construction of models for particular populations and configurations of sites and the verification of actual conservation. While it is possible to extrapolate from one landscape to another through indicator species, two sites and respective biogeographies are never alike. Conventional scientific verification and simulation has tended to stay on the margins of these questions through various forms of reductionism. These earlier conceptions for providing the basis for spatially precise conservation planning and management across regions are increasingly bankrupt, particularly where there is recognition of the benefits from modelling cumulative relationships. The various simplistic guidelines for coping with complex conservation problems are increasingly losing their currency. Aside from the geographic information system-based approaches to gap analysis, there have been few successful district-wide simulations for the conservation of biological diversity, involving both a range of natural and social variables as part of the identification of viable and socially acceptable strategies. The first response to recognition of the problems of empirical verification of landscape relationships over time for conservation simulation was the development of island biogeography theory. Too often, however, the focus of the theory has been simply on validating generalizations pointing to the need for more areas of protected habitat. Landscape ecology when linked to conservation biology theory could provide the necessary framework for monitoring and modelling such complex sets of natural and cultural attributes for simulations and the formulation of a number of alternative sets of interventions for both habitat protection and economic development. But a larger problem remains. We cannot look into the future several hundred years to monitor the outcomes of divergent conservation strategies and the postulates that they reflect in order to determine which path to take today. The current extrapolations from a few short-term trends and spatial relationships are poor substitutes for simulating complex interactions and possible conservation measures. A new kind of cumulative conservation biology is needed. Such emerging investigative frameworks could rely heavily on spatial and temporal modelling and embody less reductionist strategies for overcoming gaps in knowledge and theory for particular ecosystems. The requirements for considering both social and natural factors in spatial modelling will require new algorithms ^14, derived in part from expanded conceptions of "community ecology" ^15. The use of such cumulative models must clearly acknowledge both the provisional and adaptive nature of regional conservation strategies and the reality that both biota and conservation respond to fluid, unique and site-specific relationships. In terms of the three postulates, a four dimensional matrix can be constructed with eight generic conservation strategies ^16. Some of these strategies emphasize the discrete island approach while others more fully reflect permeability and connectivity. But these polarities are far more supple and multi-dimensional than the SLOSS (single large or several small) dichotomy (Lomolino 1994). Island strategies have tended to be associated with large park bureaucracies, with high levels of state and even military backing, while the membrane strategies are often associated with efforts to allow local communities to live within park systems and to direct ecosystem management for both conservation and social development (Ingram 1994a). The complex implications of the various ranges of conservation possibilities for social development are also worth new investigations and simulations. New indicators of the effectiveness of singular and grouped conservation measures can be meshed with those for persistence of particular elements of biodiversity. Additional postulated tradeoffs for conservation must quickly be explored in order to deepen interpretations of what little we know about and hope for the persistence of the many facets of natural and human life across landscapes. Design processes and management frameworks for the expansion of district networks of protected areas, based on these postulates, recognize the inherent instability of ecosystems across landscapes, the shifting political economies, and the erratic social patterns of aesthetics and fashions. The kinds of protected area design processes envisioned here recognize that while protected areas must become "more effective" at insuring the persistence of local biodiversity, such public lands will also be serving an increasingly intense and diverse set of social functions not directly related to biodiversity conservation. As intensifying land use and secondary and tertiary ecological disturbances effectively cast protected areas further adrift, it will be necessary to design and manage landscapes to maximize effectiveness for an uncertain future. Building moats around habitat islands was never enough. Instead, there are more precise and efficient bases for balancing a range of social needs. ACKNOWLEDGEMENTS These postulates first emerged in discussions with Jeff Romm and the late Thomas Dickert at the University of California, Berkeley in 1983 - 85. They were based on interdisciplinary field work in Niger (1994 - 96), Cameroon (1996), Yemen (1994), Indonesia (1986 1988 1989, and 1990) and the Pacific coast of Canada (1988, - 1995). In subsequent years, Bill Lidicker, Richard Meier and Robert Twiss, also of Berkeley, and Ray Dasmann of the Environmental Studies Board of the University of California at Santa Cruz, gave crucial support to these inquiries. Recently, Craig Shafer, Richard Meier, Ferdinando Villa, and former student and University of British Columbia graduate, Vince Amendolagine, contributed new perspectives on the nature of the curves of the axes of minimum tradeoffs. Funding over the last ten years has come from the CGIAR (the Consultative Group on International Agricultural Research), the University of California at Berkeley, IUCN (The World Conservation Union), FAO (the Food and Agriculture Organization of the United Nations), the British Columbia Ministry of Forests, CIDA (Canadian International Development Agency) and IDRC (International Development Research Centre of Canada). REFERENCES Athnanasiou, T. 1996. Divided Planet: The ecology of rich and poor. New York: Little, Brown and Company. (published in Europe in 1997 as Slow Reckoning: The Ecology of a Divided Planet. London: Seeker & Warburg). Barker, M. L. 1982. Comparison of parks, reserves and landscape protection in three countries of the eastern Alps. Environmental Conservation 9: 275 - 285. Barrett, T. S. and P. Livermore. 1983. The Conservation Easement in California, Island Press, Covelo, California. Buechner, M., C. Schonewald-Cox, R. Sauvajot, and B. A. Wilcox. 1992. Cross-boundary issues for national parks: What works "on the ground." Environmental Management 16: 799 - 809. de Klemm, C. 1985. Preserving genetic diversity: A legal review. Landscape Planning 12: 221 - 238. Deutsche, R. 1996. Evictions: Art and spatial politics. Cambridge, Massachusetts: MIT Press. Diaz, N. and D. Apostol. 1992. Forest Landscape Analysis and Design: A process for developing and implementing land management objectives for landscape patterns, US Department of Agriculture Forest Service, Gresham, Oregon. Diamond, J. M. 1975. The island dilemma: Lessons of modern biogeographic studies for the design of nature reserves. Biological Conservation 7: 129 - 146. Dyer, M. I. and M. M. Holland. 1991. The biosphere-reserve concept: Needs for a network design. BioScience 41: 319 - 325. Feyerabend, P. 1988. Against Method (revised edition). New York, Verso. Forman, R. T. T. and M. Godron. 1986. Landscape Ecology. New York, John Wiley & Sons. Frankel, O. H., and M. E. Soule. 1981. Conservation and Evolution. Cambridge University Press, New York. Giacomini, V. and V. Romani. 1978. National parks as open systems: An Italian overview. Landscape Planning 5: 89 - 108. Given, D. R. 1994. Principles and Practice of Plant Conservation. Portland, Oregon: Timber Press / WWF / IUCN. Grumbine, R. E. 1994. What is ecosystem management? Conservation Biology 8(1): 27 - 38. Harmon, David (editor). 1994. Coordinating Research and Management to Enhance Protected Areas. The World Conservation Union (IUCN), Cambridge, UK. Harvey, D. 1989. The Condition of Postmodernity. An Enquiry into the Origins of Cultural Change. Cambridge, Massachusetts: Blackwell. Heywood, V., G. B. Ingram, J. Black and L. Olivier. 1993. Managing protected areas to conserve genetic resources. Report of Workshop IV.6 In Parks for Life. Report on the IVth World Congress on National Parks and Protected Areas. Gland, Switzerland, The World Conservation Union. pages 175 and 176. Holling, C. S. (editor). 1978. Adaptive environmental assessment and management, John Wiley, Chichester, New York. Hoose, P. M. 1981. Buildings An Ark: Tools for the preservation of natural diversity through land protection, Island Press, Covelo, California. Ingram, G. B. 1983. Parks in the 21st Century. Not Man Apart. February / March 1983: 7. Ingram, G. B. 1984. In situ conservation of plant genetic resources: The scientific and technical basis. Rome, FAO. Ingram, G. B. 1987. Conservation of wild plants in crop gene pools and their intraspecific variation: Current needs and opportunities in the moist forest of SE Asia. In The Conservation and Management of Endangered Plants and Animals. Proceedings of the Symposium on the Conservation and Management of Endangered Plants and Animals, Bogor, Indonesia, June 1986. C. Santiapillai and K.R. Ashby (eds.) 63-85. Bogor, Indonesia, South- East Asia Center for Tropical Biology. Ingram, G. B. 1989. Planning district networks of protected habitat for conservation of biological diversity: A manual with applications for marine islands with primary rainforest. Ph. D. diss. University of California, Berkeley. on file UC Berkeley and University Microfilms International, Ann Arbor, Michigan dissertation order number 9006370. Ingram, G. B. 1990a. Multi-gene pool surveys in areas with rapid genetic erosion: An example from the Air Mountains, northern Niger. Conservation Biology (New York) 4(1): 78 - 90. Ingram, G. B. 1990b. The management of biosphere reserves for the conservation and utilization of genetic resources: The social choices. Impact of Science on Society (Paris) 158: 133 - 141. Ingram, G. B. 1990c. The need for knowledge from indigenous communities in planning networks of protected habitat for the conservation of biological diversity: Three island settings. In Ethnobiology: Implications and applications. Proceedings of the First International Congress on Ethnobiology (Belem, Brazil 1988). Part 2. M.J. Plotkin (ed.), pp. 87 - 105, Belem, Para, Goeldi Museum. Ingram, G. B. 1991. Biological, visual and recreational values and the planning of extractive development and protected areas: A tale of three islands. Landscape and Urban Planning (Amsterdam) 21: 109 - 129. Ingram, G. B. 1994. Institutional obstacles to conservation of habitat and biological diversity on Fergusson Island, Milne Bay Province, Papua New Guinea. Pacific Affairs (Vancouver) 67(1): 26 - 45.
Ingram, G. B. 1994. Rainforest conservation initiated by traditional island communities: Implications for development planning. Canadian Journal of Development Studies (Ottawa) XV(2): 193 - 218. Ingram, G. B. 1992a. Fragmentation: Towards an expanded of the vulnerability of forest habitats on islands. Proceedings of the symposium, In Harmony with Nature, International Conference on the Conservation of Tropical Biodiversity. Kheong, Y. S. and L. S. Win (editors). 94 - 121. Kuala Lumpur, Malaysia, Malayan Nature Society. Ingram, G. B. 1992b. Landscape indicators for conservation of biological diversity: An example from Haida Gwaii, British Columbia. In Landscape Approaches to Wildlife and Ecosystem Management. G. B. Ingram and M. R. Moss (editors). Morin Heights, Quebec, Polyscience. 99 - 134. Ingram, G. B. 1992c. The remaining islands with primary rainforest: A global resource. Environmental Management (Massachusetts) 16(5): 585 - 595. Issue on problems on small islands. Ingram, G. B. 1994a. Rainforest conservation initiated by traditional island communities: Implications for development planning. Canadian Journal of Development Studies XV(2): 193 - 218. Ingram, G. B. 1994b. The ecology of a conflict. In Clayoquot & Dissent. Vancouver, Ronsdale Press. pages 9 - 71. Ingram, G. B. 1994c. Institutional obstacles to conservation of habitat and biological diversity on Fergusson Island, Milne Bay Province, Papua New Guinea. Pacific Affairs (Vancouver) 67(1): 26 - 45. Ingram, G. B. 1995a. Conserving habitat and biological diversity: A study of obstacles on Gwaii Haanas, British Columbia. Forest and Conservation History (North Carolina) 39(2): 77 - 89. Ingram, G. B. 1995b. Landscapes of (un)lawful chaos: Conflicts around temperate rain forest and biological diversity in Pacific Canada. RECIEL: Review of European Community & International Environmental Law (London) 4(3): 242 - 249. Ingram, G. B. 1996. Integration of in situ conservation of genetic resources into landscape and regional planning frameworks. Pages 454 - 476 In Biodiversity in Managed Landscapes: Theory and Practice. Robert C. Szaro and David W. Johnston. New York, Oxford University Press. Ingram, G. B. and J. T. Williams. 1993. Gap analysis for in situ conservation of crop genepools: Implications of the Convention on Biological Diversity. Biodiversity Letters (London) 1: 141 - 148. Jain, S. K. 1975. Genetic reserves. In Crop Genetic Resources for Today and Tomorrow. eds. O. H. Frankel and J. G. Hawkes. pp. 379 - 396. Cambridge University Press, London. Janzen, D. H. 1983. No park is an island: Increase in interference from outside as park size decreases. Oikos 41: 402 - 410. Kuhn, T. S. 1970. The Structure of Scientific Revolutions. Chicago, University of Chicago Press. Leopold, A. S., S. A. Cain, C. M. Cottam, I. N. Gabrielson and T. L. Kimnball. 1963. Wildlife Management in the National Parks. (on file) Washington, D. C., United States Department of Interior National Park Service. Lomodino, M. V. 1994. An evaluation of alternative strategies for building networks of nature reserves. Biological Conservation 69: 243 - 249. Lusigi, W. J. 1978. Planning Human Activities on Protected Natural Ecosystems, J. Cramer, Lehre, Germany. Machlis, G. E. 1992. The contribution of sociology to biodiversity research and management. Biological Conservation 62: 161 - 170. Machlis, G. E. and D. L. Tichnell. 1985. The State of the World's Parks: An international assessment for resource management, policy, and research, Westview Press, Boulder, Colorado. Main, A. R. and M. Yadov. 1981. Conservation of macropods in reserves in western Australia. Biological Conservation 10: 13 - 21. Martinez, A. 1995. The new regional planning and implementation of the Convention on Biological Diversity. In Conservation of Biodiversity and the New Regional Planning. Richard E. Saunier and Richard A. Meganck (eds.). Washington, DC: Organization of American States / IUCN. pp. 93 - 99. Meier, R. A. 1995. Creating wildlife: synthesis of a gaming simulation. Ekistics 239 (October 1975): 291 - 297. Naveh, Z. 1994. Biodiversity and landscape management. In Biodiversity and Landscapes: A paradox of humanity. K. C. Kim and R. D. Weaver (eds.). New York: Cambridge University Press. pp. 187 - 207. Noss, R. 1990. Indicators for monitoring biodiversity: A hierarchical approach. Conservation Biology 4: 355 - 364. Noss, R. F. and L. D. Harris. 1986. Nodes, networks, and MUMs: Preserving diversity at all scales. Environmental Management 10: 299 - 309. (MAB) Man and the Biosphere Programme (of Unesco). 1974). Criteria and Guidelines for the Choice and Establishment of Biosphere Reserves, Unesco, Paris. Possiel, W., R. E. Saunier, and R. A. Meganck. 1995. In-situ conservation of biodiversity. In Conservation of Biodiversity and the New Regional Planning. Richard E. Saunier and Richard A. Meganck (eds.). Washington, DC: Organization of American States / IUCN. pp. 7 - 20. Pyle, R. M. 1980. Management of nature reserves. In Conservation Biology: An evolutionary perspective, ed. M. E. Soule & B. A. Wilcox, Sunderland, Massachusetts, Sinauer Associates, pp. 319 - 328. Roberts, L. 1988. Hard choices ahead on biodiversity. Science 241: 1759 - 1761. Schonewald-Cox, C. M. and J. W. Bayless. 1986. The boundary model: A geographical analysis of design and conservation of nature reserves. Biological Conservation 38: 305 - 322. Schonewald-Cox, C., M. Buechner, R. Sauvajot, B. A. Wilcox. 1992. Environmental auditing: Cross-boundary management between national parks and surrounding lands: A review and discussion. Environmental Management 16: 273 - 282. Scott, J. M., F. Davis, B. Csuti, R. Noss, B. Butterfield, C. Groves, H. Anderson, S. Caicco, F. D'Erchia, T. C. Edwards, J. Ulliman, and R. G. Wright. 1993. Gap Analysis: A geographic approach to protection of biological diversity. Supplement to the Journal of Wildlife Management 57(1), Wildlife Monographs 123: 1 - 41. Shafer, C. 1994. Beyond park boundaries. In Landscape Planning and Ecological Networks. E. A. Cook and H. N van Lier (eds.). Amsterdam: Elsevier. pp. 201 - 223. Shaffer, M. L. 1987. Minimum viable populations: coping with uncertainty. In M. Soule (ed.). Viable Population for Conservation. London: Cambridge University Press. 69 - 86. Slayter, R. O. 1974. A National System of Ecological Reserves in Australia, Canberra, Australia. Slocombe, D. S. 1995. An ecosystem approach to regional planning. In Conservation of Biodiversity and the New Regional Planning. Richard E. Saunier and Richard A. Meganck (eds.). Washington, DC: Organization of American States / IUCN. pp. 53 - 66. Soule, M. E. and G. Lease (eds.). 1995. Reinventing Nature? Responses to postmodern deconstruction. Washington, DC: Island Press. The Crucible Group. 1994. People, Plants, and Patents: The Impact of Intellectual Property on Biodiversity, Conservation, Trade, and Rural Society. International Development Reserve Centre, Ottawa. UNEP (United Nation Environment Programme). 1992. Convention on Biological Diversity 5 June 1992. version: Na. 92-8314. on file, Environmental Law and Institutions Programme Activity Centre, UNEP, Nairobi. Vogel, J. H. 1994. Genes For Sale. New York, Oxford University Press. Vogel, J. H. and G. B. Ingram. 1993. Biodiversity versus `genetically coded functions': The importance of definitions in conservation policy. RECIEL: Review of European Community & International Environmental Law London) 2(2): 121 - 125. WCMC (World Conservation Monitoring Centre). 1992. In situ conservation of crops and wild relatives of crops. In Global Biodiversity: Status of the Earth's Living Resources. B. Groombridge (editor). 544 - 549. London, Chapman and Hall. WRI (World Resources Institute), IUCN (World Conservation Union), and UNEP (United Nations Environment Programme). 1992. Global Biodiversity Strategy. Washington, DC: WRI.
NOTES 1. The most important paper to establish the "island biogeography paradigm" in conservation planning was Diamond's 1995 essay "The island dilemma..." For a romanticized chronicle of the emergence of the key ideas of island biogeography and conservation biology, see David Quammen's 1996 The Song of the Dodo: Island biogeography in an age of extinctions. 2. Much of my development of a notion of archipelagos has been based on the work of Jared Diamond and his kind encouragement in the period from 1983 to 1990. 3. There are a myriad of definitions and interpretations of "biological diversity" and "biodiversity". I suggest the reader picks three articles or books randomly from conservation publications over the last decade, and focus on the differences in those interpretations. Such comparison will yield substantial differences that have direct bearings on specific conservation and land management objectives. 4. This paper is being reworked in the upcoming book by G. B. Ingram and F. Villa, "Tradeoffs in Conservation: Alternative Scenarios for Planning Networks of Open Space and Protected Areas." 5. The notion of trade-off as for identification of a variable outcome involving similar level of environmental conservation, has been a theme for over twenty years. The term "tradeoffs" has been used in many different ways in land management. For a discussion of a contrasting use of the word "trade-offs," employed in justifying lowering environmental standards, see Athnanasiou (1996) p. 187. 6. In using the notion of "simulation" in predictions of biodiversity conservation - activities that today are nearly always computerized and spatialized, I am influenced by Richard Meier's (1995) notion of "gaming." 7. Some of the more recent and overt framings of modernist generalizations in conservation biology were made in Michael E. Soul and Gary Lease (eds.) (1995). 8. The most influential single treatment that argued for postmodernity was Donald Harvey's 1989 'The Condition of Postmodernity. An Enquiry into the Origins of Cultural Change', though in recent years there have been various challenges to it, most notably Rosalyn Deutsche's 1996 'Evictions: Art and spatial politics' with her scathing critique, "Boys Town". 9. For a review of the uses of "site-specificity" as a cultural movement, but with direct implication for scientific investigations and environmental management, see Rosalyn Deutsche. 1996. Evictions: Art and spatial politics, pp. 92-93, 140. 10. There have been numerous minor revisions and programme documents since the 1992 document that was signed at the UNCED meeting in Rio de Janeiro in 1992. I cite this version because it was probably the truest to the original compromise that was embodied in the Convention of Biological Diversity - with all of its contradictions. 11. While "regulation" often suggests state intervention, I use it in a broader sense, as in some kind of social restraint system for conservation. This in turn includes state- sanctioned prohibitions as well as less adversarial "cooperative" (Shafer 1994) arrangements involving various non-governmental, private and more public sectors. 12. The conflicts over protected area designs that has most influenced my perspectives in recent years have been around rain forest on islands (Ingram 1991, 1992c, 1994a, 1994c, 1995b), around the establishment of Gwaii Haanas National Park (Ingram 1995a) on the Queen Charlotte Islands of Pacific Canada and around Clayoquot Sound on Vancouver Island (Ingram 1994b). 13. Shafer (1994, p. 223) cites this quotation as from "Wright, G.M. and B.H. Thompson, 1935. Fauna of the National Parks of the United States: Wildlife Management in the National Parks. Fauna Series No. 2. U.S. Government Printing Office, Washington, D.C.). 14. In 1997, there are numerous geographic information systems algorithms available, especially in the forms of macros, "aml"s and more extensive software modules, for interactive spatial exploration of options for conservation and land use. Much of this work has a connection to the gaming theory of the 1960s and 1970s (pers. comm. Prof. Richard Meier, College of Environmental Design, University of California Berkeley - email: meier@ced.berkeley.edu). "Nowadays I would, like you, get the stakeholders together to explore the options list, reduce the situation to a GO board and scale, populations to 5-9 indicator species, reproduce historic catastrophes, and interventions...It is most important to keep everything transparent when proceeding through time. This model should be subjected to scenarios by the stakeholders, and readjusted to satisfy them. Then GIS data can be superimposed, and socio-political events at different stages of the scenarios, to discover the range of impacts... Long ago I wrote a paper about the process. Maybe I can find a copy, but it had little impact. I think yours could, if it introduced simulation synthesis, because the mangers, planners, and designers are quickly moving in that direction, and making many expensive mistakes." (letter from Richard Meier to Brent Ingram, October 6, 1996). 15. I remain indebted to Dick Meier and Ray Dasmann for their divergent visions of "community ecology" - both approaches based on humanistic and holistic approaches far less reductionist than how the term has often been employed. 16. The eight generic strategies for planning networks of protected areas were explored in my 1989 thesis, and is the focus of several chapters in the upcoming book by G. Ingram and F. Villa, "Tradeoffs in Conservation: Alternative scenarios for planning networks of open space and protected areas." Copyright remains with the author.
Published by Bioline Publications.
The following images related to this document are available:Line drawing images[py97003b.gif] [py97003a.gif] [py97003c.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}