 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
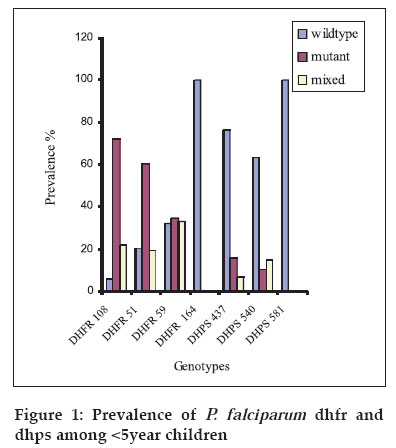
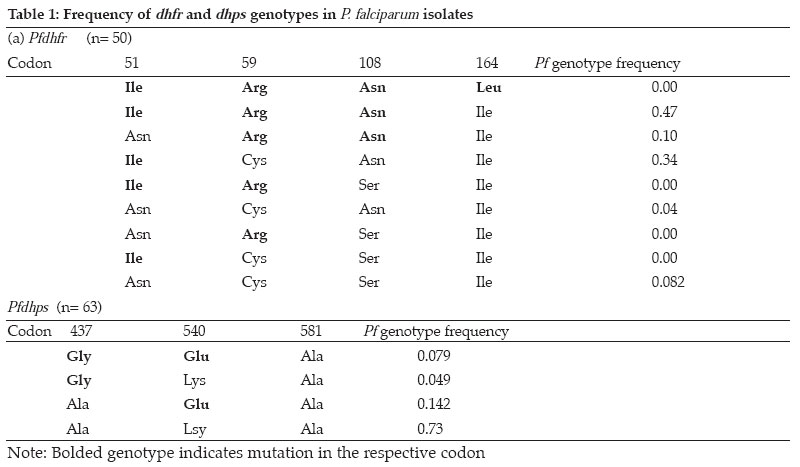
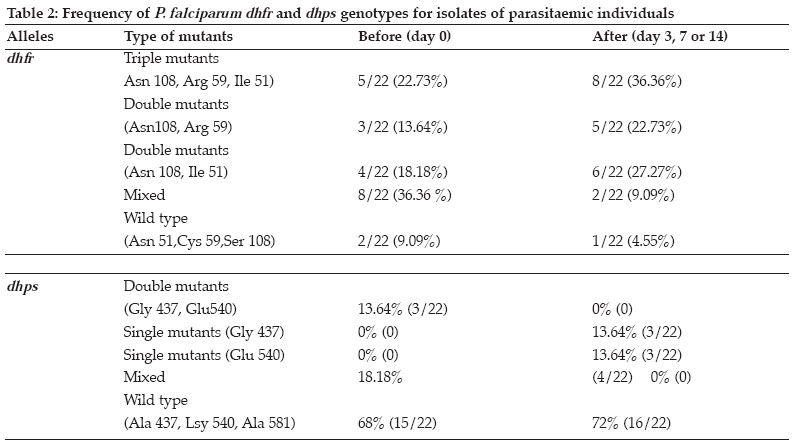
Tanzania Health Research Bulletin, Vol. 8, No. 2, May, 2006 pp. 50-55 Dhfr and dhps mutations in Plasmodium falciparum isolates in Mlandizi, Kibaha, Tanzania: association with clinical outcome W. KIDIMA1*, G. NKWENGULILA1, Z. PREMJI2, A. MALISA3 and H. MSHINDA3 1University of Dar-es-Salaam, Department of Zoology and Wildlife Management, P. O. Box 35064, Dar-es-Salaam, Tanzania * Correspondence : W. Kidima; E-mail: winikid@yahoo.co.uk Code Number: rb06011 Abstract: Sulfadoxine-pyrimethamine (SP), the current first line antimalarial drug in Tanzania, is compromised by evolution and spread of mutations in the parasite's dhfr and dhps genes. In the present study we established the baseline frequencies of Plasmodium falciparum dihydrofolate reductase (pfdhfr) and dihydropteroate synthase (pfdhps) mutant genotypes and their potential for predicting the in vivo efficacy of SP in Mlandizi, Tanzania. The efficacy of SP treatment was by following 116 children with uncomplicated falciparum malaria for 14 days after treatment. Infected blood samples were collected on filter paper at days 0, 3, 7 and 14. Parasite genomic DNA was extracted and point mutations at positions 51, 59, 108 and 164 of the dhfr gene and at 581, 540 and 437 of the dhps gene were analysed by nested Polymerase Chain Reaction/ Restriction Fragment Length Polymorphism. Out of 116 children enrolled, 98 (86%) of eligible children demonstrated an adequate clinical response by day 14. There were 7.3 % early and 6.7% late therapeutic failures. At day 0, only 8.0% (4/50) the parasites showed no mutation at the dhfr locus; for dhps this was 73%. Triple mutant dhfr alleles (Ile 51, Arg 59, Asn 108) occurred in 47%, double mutant dhps (Gly 437, Glu 540) alleles in 7.9%. No mutation was detected at codon 164 of the dhfr gene. The presence of triple dhfr mutant alleles was related to clinical failure, but did not show significant association (Fisher exact test, P=0.166, OR 2.15 0.77 Key words: Plasmodium falciparum, sulfadoxine-pyrimethamine, dhfr, dhps, Tanzania Introduction Malaria parasites have resisted all attempts to be eradicated, either by drug treatment to kill the parasites themselves, or by elimination of the mosquito vectors. The reason for this failure is complex, but stems partly from the fact that the parasites are genetically complex organisms with a powerful ability to generate mutants that, for example, confer resistance to drugs (Walliker et al., 1998). Drug resistance in malaria parasites is a serious problem in Plasmodium falciparum (Greenwood & Mutabingwa, 2002). Chloroquine has been rendered ineffective by resistant parasites in many parts of the world, the emergence of which has been linked to an increase in childhood morbidity and mortality in African children (Bloland, 2001). A fixed combination of sulfadoxine-pyrimethamine (SP), the current first line antimalarial drug in Tanzania and other Southern African countries, may be compromised by the rapid emergence and spread of resistance. This resistance emerges as a result of mutations on the dihydrofolate reductase (dhfr) and dihydropteroate synthase (dhps) enzyme systems of P. falciparum. In Africa, resistance to SP is associated with three mutations in the dhfr gene (Ile 51, Arg 59, Asn 108 ) and two mutations in the dhps gene (Gly 437, Glu 540 ) (Kublin et al., 2002). Molecular markers are potential indicators to the deteriorating efficacy of an antimalarial drug. In longitudinal studies, pfdhfr and pfdhps markers were used to determine antifolate resistance by P. falciparum (Nzila et al., 2000). The identification and implications of molecular markers for antimalarial drug resistance is regarded key to elucidating and eventually controlling resistance (Wernsdorfer & Noedl, 2003). Molecular markers can provide direct and convincing evidence of selection of resistant parasites (Nzila et al., 2000; Doumbo et al., 2000). However, whether the presence of resistance also predicts the outcome of treatment is less clear. In the present study the potential of pfdhfr and pfdhps mutant genotypes to predict the clinical response to SP treatment among children was evaluated. The hypotheses were that the presence of triple pfdhfr mutant genotypes and double mutant pfdhps in an individual before treatment predicts persistent infection and that SP would select for pfdhfr and pfdhps mutations during a short period of treatment. Only mutations at codons 51, 59, 108 and 164 of dhfr gene and 437, 540 and 581 of dhps gene were investigated. Materials and Methods Study area and population The study was carried out in Mlandizi Township in Kibaha district, Tanzania, as part of the National Malaria Control Programme to monitor antimalarial drug resistance. Mlandizi was an ideal location for conducting such a programme, as malaria transmission is stable and intense for most of the year with a mean annual entomological inoculation rate of over 200 infective bites per person per year (Premji et al., 1997). Children aged 1-59 months attending Mlandizi Health Centre were recruited for the survey during the 2002 rainy season. Children presenting with non-severe malaria attack were enrolled in the survey. Inclusion criteria were monoinfection with P. falciparum with at least 300 parasites/µl, no intake of antimalarials or sulphonamide-based drugs during the previous 4 weeks, absence of severe illness, presence of axillary temperature of ≥ 37.5°C but <39.5°, no history of allergic reactions to the administered drug and willingness to give consent to participate. Patients were treated (under observation) with a standard dose of SP (Fansidar® Roche), 1.25mg/kg of pyrimethamine and 25mg/kg of sulfadoxine. Upon presentation to day 0 and during follow up visits on day 3, 7 and 14, thick and thin blood smears were collected by finger prick and stained with Giemsa for microscopic parasite identification and quantification. In the thick smear parasites were counted against 200 white blood cell count, assuming this to be 8000/µl. The parasites density was expressed as to the number of parasites/µl. Assessment of the treatment response was based on the WHO 14 day's in vivo protocol (WHO, 1996). At enrolment and during the follow-up visits, finger prick blood was also spotted on filter paper (3mm Whatmann International Ltd. Maidstone, United Kingdom). Filter papers were air-dried and stored in separate envelopes at room temperature until required for DNA extraction. Parasite genomic DNA was extracted by chelex extraction as described elsewhere (Plowe et al., 1995). Drug resistance markers point mutation analysis Parasite genomic DNA was analysed for SP resistance markers on the dhfr (chromosome 4) and dhps (chromosome 8) genes by using a nested Polymerase Chain Reaction/ Restriction Fragment Length Polymorphism (PCR/RFLP) as described by Duraisign et al. (1998). Mutations on pfdhfr codons 51, 59, 108 and 164 and pfdhps codons 437, 540 and 581 were determined. The primary reaction involved a multiplex PCR. Nested reaction for dhfr and dhps were carried out separately for each inner primer pairs that were used. The nested PCR products were monitored on 2% agarose gel (MetaPhorTM USA) stained with ethidium bromide prior to restriction digestion and photographing. Various laboratory strains of P. falciparum were used as positive controls in the PCR reaction, including 3D7, FCR3, V1S, W2, T9/96, HB3 and K1 (MR4). A tube without DNA template was also included in the reaction as a negative control. Positive PCR products were genotyped by RFLP analysis as previously described (Duraisign et al., 1998). Restriction digest products were then run on 10% polyacraliamide gel (Q-Bio gene, Canada) using 1.5mm spacers, stained by 0.5µg/ml of ethidium bromide, viewed under UV light and photographed using Polaroid and scored. P. falciparum isolates from treatment failures were genotyped by PCR analysis targeting merozoite surface protein (MSP2) gene. The method included a nested PCR followed by analysis by RFLP (Felger et. al., 1999). Since the parasite is haploid in human host, changes of the respective parasite genotypes after infection indicates a new infection, whereas detection of the same parasite genotype indicates a recrudescent parasite. Ethical approval was given by the Tanzania Medical Research Coordinating Committee (Ref. No. NIMR/HQ/R.8a /Vol.1X / 107). Verbal informed consent was obtained from village leaders and written consent was obtained from all parents or guardians prior to enrolment in the survey. Data analysis Data was analysed using Epi-Info-6. Prevalence of each mutation was expressed as the ratio with the total number of isolated infections. The baseline mutation rates were determined from sample taken on day 0. Chi-square test was used to compare prevalence rates of triple pfdhfr mutant genotype and double pfdhps mutant genotypes. Fisher' exact test was used to analyze the association between different dhfr and dhps genotypes and treatment failures. Mixed infections were excluded from statistical analysis. All the probabilities are two tailed and results recorded as significant at P<0.05. Results Treatment outcome and frequency of dhfr and dhps genotypes in P. falciparum isolates Out of 116 enrolled children, 2 (2%) were lost to follow up. Adequate clinical response was observed among 99 (86%) children. Early and late treatment failures were observed in 8 (7.3%) and 7 (6.7%) children, respectively. Out of 114 samples only 106 were successfully genotyped. Mutations in the dhfr gene were more prevalent than those of dhps gene (Figure 1). Most frequently dhfr alleles contained mutant at one or more codons studied and few were wild types. At dhfr locus the commonest mutant alleles were Asn 108 / Arg 59/ Ile 51 and Asn 108/ Ile 51 (Table 1), a combination of dhfr allele reported to confer resistance to SP. At the dhps locus however the most frequent allele was the wild type form, Ala 437/ Lys 540/ Ala 581. The frequency of occurrence of triple pfdhfr (Asn 108, Arg 59 and Ile 51) mutation was more prevalent than that of double pfdhps (Gly 437, Glu 540) mutation ( χ2 =12.004, P= 0.0005). Dhfr and dhps genotypes in relation to treatment outcome Results of the genetic analysis of the isolates before and after treatment with SP failures are summarised in Tables 2. A parasite with a quintuple mutation (Asn 108, Arg 59, Ile 51, Glu 437 and Gly 540) was observed in only one isolate that demonstrated a late treatment failure. The analysis of in vivo failures of recrudescent single infections showed no association between presence of triple pfdhfr (Asn 108/Arg 59/Ile 51) mutation in the dhfr gene and clinical outcome (Fisher exact test, P=0.1533, OR 2.53 0.64>OR<9.74). Evolution of parasites after treatment Changes of parasite genotypes following treatment with SP were observed in most isolates. Most mixed genotypes changed to mutant genotypes, indicating selection for mutation against sensitive forms (wild type) of the parasites. There was a sufficient evidence to suggest that drug selection of more resistant subpopulations was occurring between day 0 and day 14. Among the post-treatment infections, 64.4% (14/22) of the parasite isolates were recrudescent and 35.6 % (8/22) were new infections. Discussion The present study assessed the resistance status of P. falciparum population in Mlandizi for SP, using the in vivo and molecular data. The infections of P. falciparum in Mlandizi contained many mixed parasite population with both wild type and mutant parasites. This is in agreement with the multiclonal nature of natural infections (Babiker et al., 1994; Arez et al., 1997; Farnert, 2000) in relatively high transmission areas. The occurrence of mixed infections poses a problem in estimating the proportion of mutation in each parasite clone. The genetic changes observed in the dhfr and dhps genes in this parasite population have an impact on the therapeutic response to SP drug. The high frequency of triple dhfr allele (Asn 108, Arg 59 and Ile 51) and the dhfr Asn 108 and Ile 51 combinations, suggests substantial use of pyrimethamine or related drugs in Mlandizi. These results suggest that resistance against SP have already been established and is more likely to intensify as drug pressure increases, in the near future. The observed high cure rate with SP is likely to have been attributed to the shorter day follow up (14 days). Owing to the prophylactic nature of SP, more treatment failures tend to occur after day 14. Plowe et al. (2004) observed a trend towards diminishing clinical and parasitological efficacy in 28 days follow up as compared to the 14 days. High recrudescent rates at day 28 have been reported from a study carried out in Zanzibar using long-acting drugs (Martensson et al., 2005). It is clear that the 14-day protocol has a high chance of underestimating treatment failures. Thus, in addition to the small number of samples in this study, the 14-day protocol is likely to have contributed to the insignificant association of molecular markers to treatment outcome. In the present study, there were no parasites that had quadruple mutation (Ile 51, Arg 59, Asn 108, and Leu 164). Similar findings have been reported in northern (Pearce et al., 2003) and southeastern Tanzania (Mshinda, 2000) and in Malawi (Kublin et al., 2002; Bwijo et al., 2003). On the Thai-Myanmar border, Thailand, where high levels of resistance to SP have been reported, the Ile 51/ Arg 59/ Asn 108/ Leu 164 alleles of dhfr were very common (Nair et al., 2002). The absence of Leu 164 mutation in the present study indicates that it is rare and this drug has not reached its highest level. Seemingly, the parasites with this mutation do not have a survival advantages in many parts of Africa. Reports from Sibley et al. (1997) and Nzila et al. (2005), suggest that the quadruple mutant dhfr may not be an efficient enzyme and does not confer a biological advantage in the context of the cell compared to the triple mutant (Nzila et al., 2005). The pre-treatment infections of 34 isolates presenting triple mutations on the dhfr gene did not lead to treatment failures. Either, 9 parasites demonstrating double mutation at the dhps loci 437 and 540 with either double or single mutation at dhfr loci did not influence treatment outcome with SP. These parasites might have been insensitive to SP treatment. Such inconsistencies have also been observed in a longitudinal drug efficacy study in Gabon (Aubouy et al., 2003). The inconsistencies, therefore, compromise the predictive value of molecular markers. The above observations corroborate the contention that, effective treatment is the result of a complex interaction between parasite, host and medicament (Wernsdorfer, 1994). There are other factors, other than mutation, that contribute to variability in response to a drug, including the drug level achieved by the host, parasite resistance to drug, host immune response and possibly serum levels of folate. Human folate levels can increase resistance to SP by selection of a subpopulation of parasites after drug administration (Wang et al., 1997). These authors have reported that folate supplementation is associated with SP failure as the susceptibility of pyrimethamine and sulfadoxine is compromised. In conclusion, the continued use of SP will inevitably lead to increase in drug pressure, which in turn will lead to the emergence of resistant strains of P. falciparum as mutation on both dhfr and the dhps increases. As resistance against SP increases, combination therapy may be the best option to retard its spread. The lack of significant association between the presence of molecular markers and treatment failures with SP indicates resistance against SP is a function of more factors than just mutations. Acknowledgements We would like to give sincere thanks to Christopher Membi and the molecular parasitology group at Ifakara Health Research and Development Centre for their technical assistance. This study received financial assistance from DAAD. References
Copyright 2006 - Health User's Trust Fund (HRUTF) The following images related to this document are available:Photo images[rb06011f1.jpg] [rb06011t1.jpg] [rb06011t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}