 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Revista Colombia Médica, Vol. 33, No. 2, 2002, pp. 52-57 Construcción y expresión de una proteína recombinante de la envoltura del virus linfotrópico humano tipo I (HTLV-I) Paula Ordóñez, Quim.1, Flavio Cerón2, Felipe García-Vallejo, Ph.D.3
Code Number: rc02011 RESUMEN El virus linfotrópico humano tipo I de las células T (HTLV-I) es un oncorretrovirus asociado con la leucemia de células T adultas (ATL) y la Paraparesia Espástica Tropical (PET). El gen env del virus codifica la glicoproteína precursora gp62, que mediante procesamiento produce la gp21 (transmembranal) y la gp46 (superficie). Se ha demostrado la alta inmunorreactividad de la gp46 en sueros de individuos infectados. Con el propósito de obtener una proteína recombinante útil en diagnóstico y en una potencial vacuna, se clonó en el vector de expresión de Baculovirus “bácmido” pBlueBac4.5His el segmento de 144bp obtenido mediante reacción en cadena de la polimerasa (PCR) con cebadores que amplifican la región 5622-5765 nt. El construido pBlueBacRENV fue transfectado en células del insecto Mamestrae brassicae (clon MB 8003) recuperándose la proteína recombinante. La proteína RENV reaccionó específicamente con suero de personas infectadas con HTLV-I. Los resultados obtenidos mostraron que la proteína RENV puede ser utilizada para el diagnóstico serológico de la infección por HTLV-I y en la evaluación de nuevas vacunas. Palabras clave: ADN recombinante. Retrovirus. gp46. HTLV-I. Clonación. Baculovirus. SUMMARY Human T-Cell Lymphotropic Virus Type I (HTLV-I) is an oncoretrovirus associated with Adult T-Cell Leukemia/Lymphoma (ATL) and Tropical Spastic Paraparesis (TSP). The viral env gen encodes gp62 glycoprotein precursor that is cleavage to produce the gp21 (transmembrane) and gp46 (surface). Previous reports showed that gp46 is highly immunoreactive in sera from infected individuals. With the purpose to obtain a recombinant protein to be used for diagnosis and potential vaccine, we cloned into Baculovirus expression vector pBluebac4.5His bacmids, the 144bp segment obtained by PCR with primers for 5622-5765 nt region. The construct pBlueBacRENV was transfected into insect cells of Mamestrae brassicae (MB 8003) and the recombinant protein was recovered. The protein showed cross reactions with sera of HTLV-I infected people exclusively. These results enabled us to propose this protein can be used in serologic diagnosis for HTLV-I infection and as a candidate for a vaccine. Key word: Recombinant DNA. Retroviruses. gp46. HTLV-I. Cloning. Baculoviruses. El virus linfotrópico humano tipo I de las células T (HTLV-I) presenta una elevada seroprevalencia en el suroccidente colombiano, es el agente etiológico asociado con la leucemia de células T del adulto (ATL) y con una enfermedad neurodegenerativa denominada paraparesia espástica tropical/mielopatía asociada con el HTLV-I (PET/HAM)1. La PET/HAM y la ATL son dos enfermedades completamente diferentes a pesar de estar asociadas con el HTLV-I2,3. El genoma del HTLV-I posee tres regiones genéticas que codifican los precursores de la nucleocápside (gag) del complejo transcriptasa reversa-polimerasa-integrasa (pol) y de las glicoproteínas de la envoltura viral (env). Además posee dos regiones de repetición largas en sus extremos denominadas LTRs. El producto del gen env del HTLV-I es la glicoproteína precursora gp62, que es procesada para producir las dos proteínas maduras de la envoltura del virión: la glicoproteína gp46 de superficie y la transmembranal gp21. La gp46 es esencial en las etapas tempranas de la infección viral y es la proteína más inmunogénica de todos los antígenos virales. Inicialmente Palker4, demostró que la proteína gp46 de la envoltura del HTLV-I es el antígeno viral predominante reconocido por anticuerpos circulantes en el suero de pacientes infectados con HTLV-I. También se ha demostrado que en otras infecciones retrovirales como en el caso del virus de la inmunodeficiencia humana tipo 1 (VIH-1), la mayoría de los anticuerpos neutralizantes son dirigidos contra proteínas de envoltura2. Estudios previos realizados por Palker4, revelaron variaciones en la reactividad de diferentes regiones de la proteína de envoltura del HTLV-I. Estos autores ensayaron tres péptidos sintéticos que cubrían diferentes dominios de la proteína gp62 y encontraron que la región más inmunoreactiva es la parte media que corresponde al dominio de superficie SU de la gp46. La información sobre los niveles de reactividad de anticuerpos humanos a regiones específicas de la gp62 es de gran utilidad en los procedimientos de diagnóstico clínico, tamizaje y en el desarrollo de una posible vacuna contra el HTLV-I5. Uno de los objetivos más importantes de la biotecnología molecular es lograr la expresión de la información genética en sistemas heterólogos. Uno de los sistemas más utilizados para expresar proteínas de genes clonados es el sistema de Baculovirus. Este virus posee un promotor de expresión fuerte, derivado del promotor de polihedrina, el cual se ha insertado en plásmidos bacterianos para proveer un sistema de clonación de genes eucarióticos que pueden expresarse en células de insecto. Una de las ventajas de este sistema es poder expresar proteínas virales de otros organismos en un sistema seguro como son las células de insecto. En este trabajo se construyó el vector recombinante pBlueBacRENV y se expresó en células del insecto Mamestrae brassicae (MB8003) una proteína recombinante del epítope B (161-206 a.a.) de la gp46 de la superficie del HTLV-I. El producto obtenido mostró reactividad inmunológica con el suero de pacientes infectados con el HTLV-I. Los resultados obtenidos demostraron que esta proteína recombinante puede ser potencialmente útil para el desarrollo de pruebas de detección de anticuerpos específicos para el virus. Además, se considera que esta proteína podría ser utilizada como inmunoprotectora en un potencial desarrollo de una vacuna efectiva que controle la infección por el HTLV-I. MATERIALES Y MÉTODOS El ácido desoxirribonucleico (ADN) proviral se extrajo de las células MT4 provenientes de un paciente de Japón con ATL. El procedimiento de extracción se llevó a cabo siguiendo la metodología desarrollada por Soto-Ramírez et al.6 con algunas modificaciones introducidas en el laboratorio. La pureza de las preparaciones de ADN se valoró midiendo la absorbancia a 260 nm y 280 nm. Relaciones de A260/A280 >1.8 se consideraron óptimas para reacción en cadena de polimerasa (PCR). PCR en la región env. Se realizó un PCR anidado utilizando los oligonucleótidos cebadores que aparecen en el Cuadro 1. Las condiciones de PCR para generar un amplificado de 1440 bp del gen env completo con los oligonucleótidos cebadores ENV1+ y ENV3- fueron las siguientes: un paso de 3 minutos a 94°C, 35 ciclos de 1 minuto a 94°C, 30 segundos a 48°C, 45 segundos a 72°C, un ciclo de extensión de 10 minutos a 72°C. Luego se efectuó otro PCR empleando los oligonucleótidos cebadores ENV2+ y ENV4-, para ello se emplearon las mismas condiciones del PCR anterior. El producto de este segundo PCR fue un amplicón de 1440 bp que incluye todo el gen env (5180 a 6619 bp). El fragmento de 1440 bp fue sometido a un segundo PCR empleando el par de oligonucleótidos cebadores RENV1+ y RENV2- (Cuadro 1). Para este nuevo PCR las condiciones fueron: un paso de 3 minutos a 94°C, 35 ciclos de 1 minuto a 94°C, 30 segundos a 45°C, 45 segundos a 72°C y un ciclo de extensión de 10 minutos a 72°C. Después se realizó una electroforesis en gel de agarosa 2% del fragmento obtenido cuyo tamaño fue de 144 bp. Con el fin de recuperar los amplificados se realizó la electroforesis en el gel de agarosa de bajo punto de fusión y se empleó el estuche QiaGen® de elución de bandas siguiendo los procedimientos recomendados por los fabricantes. Oligonucleótidos cebadores empleados en la obtención del gen env completo y el segmento recombinante RENV
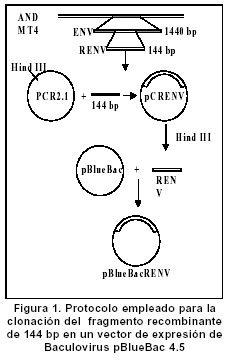
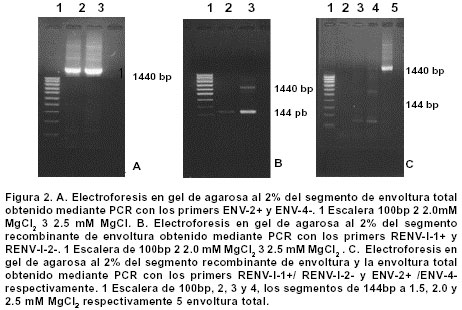
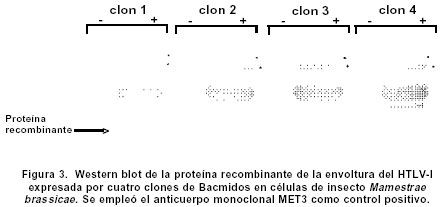
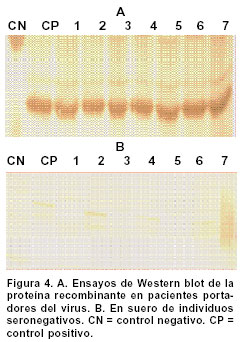
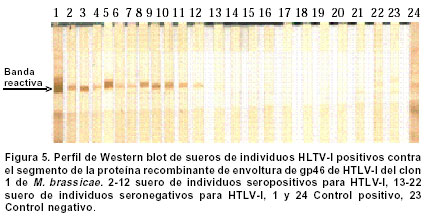
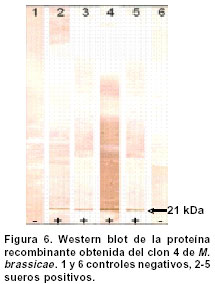
Experimentos de clonación.El fragmento de 144 bp fue clonado en el sitio Hind III del vector pCR2.1. Este vector se empleó como control de clonación del amplificado que luego se insertó en un vector de Baculovirus. La reacción de ligación se realizó a partir de los amplificados obtenidos en la elución de bandas y el vector pCR2.1 (Invitrogen®) digerido con Hind III; luego se repitió el proceso con el vector de Baculovirus pBlueBac4.5 (Figura 1). Este vector de transfección de Baculovirus se utilizó para reemplazar el gen de polihedrina tipo AcMNPV por un gen heterólogo, en este caso el gen que codifica para el fragmento de 144 bp de la gp46 de HTLV-I. Al vector recombinante obtenido se le denominó pBlueBacRENV. Secuenciación de ácidos nucleicos. Para la secuenciación del fragmento de 144 bp, se utilizó el estuche “Big Dye Terminator” (QiaGen®), siguiendo las recomendaciones de los fabricantes. La secuenciación se realizó mediante la metodología de secuenciación cíclica utilizando un secuenciador automático AbiPrism serie 3700. Expresión de la proteína recombinante. La expresión de la proteína recombinante se logró por transfección del construido pBlueBac RENV en células de insecto. Se empleó el protocolo de transfección con fosfato cálcico7 que es eficiente para la introducción de genes clonados en plásmidos recombinantes. Se colectaron por centrifugación células 3x106 de Mamestrae brassicae (MB3008) (obtenidas de la Fundación In Vitro) que se utilizaron para los experimentos de transfección. Después, a partir de lisados celulares, se purificó la proteína mediante cromatografía de ultra filtración en Sephadex G-50 hasta obtener una sola banda de Western blot8. ELISA de la proteína recombinante. Se depositaron cantidades variables de proteína recombinante (50 a 100 mg) en el fondo de los 96 pozos de una microplaca para ELISA; ésta se incubó durante toda la noche a 4ºC. Posteriormente se bloqueó con una solución de 5% de albúmina bovina fracción V (BSA) diluida en solución fosfato salina (PBS). A continuación se incubaron tanto sueros como los anticuerpos monoclonales MET3 y LAT27 para la proteína de la envoltura gp46 (donados por el Dr. Y. Tanaka) con la proteína recombinante RENV y se procedió a efectuar la prueba de ELISA9. Los sueros de los individuos incluidos en el estudio diluidos 1:100 y los anticuerpos monoclonales 1:500 se incubaron con la proteína recombinante RENV durante toda la noche a 4°C. Después de tres lavados en PBS pH7.0 durante 5 minutos cada uno, se procedió a incubar a 37°C con el anticuerpo secundario humano acoplado con streptavidina-peroxidasa durante una hora. Después de este proceso se lavó 3 veces durante 5 minutos en PBS pH7.0; la reacción se reveló mediante adición de biotina, diaminobencidina y peróxido de hidrógeno. La densidad óptica (DO) se calculó mediante un microlector Spectra II a una longitud de onda de 450 nm. Para determinar el punto de corte de la prueba Elisa se tomó el promedio de tres lecturas de sueros negativos al cual se le adicionó 1.5 veces el valor de la desviación estándar. Todas aquellas lecturas que mostraron valores mayores al punto de corte se consideraron como reactivas. Se utilizaron un total de 75 sueros de personas infectadas con el virus y 75 sueros de sujetos seronegativos tanto para el HTLV-I como para el VIH-1. Cálculos estadísticos. Las diferencias de reactividad entre sueros negativos y positivos para el HTLV-I se evaluaron mediante la aplicación de la prueba exacta de Fisher. RESULTADOS Inicialmente se realizó la amplificación con el par de primers ENV-1+ y ENV-3- y luego con el segundo par de primers ENV-2+ y ENV-4-, lo cual ofreció mayor especificidad en la amplificación (Figura 2A). Se obtuvo el fragmento recombinante de 144bp a partir de PCR (Figura 2 B,C). La secuencia de nucleótidos del fragmento amplificado y clonado (5625-5762 nt) fue: TTT TAC TCA AAA AGT TTC ACG CCT CAA TAT TAA TCT CCA TTT TTC AAA ATG CGG TTT TCC CTT CTC CCT TCT AGT CGA CGC TCC AGG ATA TGA CCC CAT CTG GTT CCT TAA TAC CGA ACC CAG CCA ACT GCC TCC CAC CGC CCC. Esta secuencia representó 100% del segmento del provirus MT4. Mediante el protocolo empleado en la elución de bandas del fragmento recombinante de 144bp, se logró obtener los amplificados para su utilización en la ligación con el sistema de Baculovirus pBlueBac4.5. Mediante traducción simulada empleando el código genético nuclear, se obtuvo la secuencia de aminoácidos del dominio de la superficie de envoltura gp46 recombinante clonada en el bacmido: Para comprobar la reactividad de la proteína recombinante obtenida, se realizaron ensayos de Western blot con sueros referencia positivos y negativos para HTLV-I. Se obtuvieron las proteínas recombinantes de células de M. brassicaey de E. coli cuyas reactividades inmunológicas fueron ensayadas con anticuerpos monoclonales para la envoltura (LAT27 y MET3). Inicialmente se observó una banda de inducción que reaccionó específicamente con los monoclonales de la envoltura (Figura 3). Varios clones recombinantes de M. brassicae se utilizaron para determinar la reactividad del anticuerpo monoclonal MET3, observándose una sola banda reactiva que no se visualizó con los sueros de individuos seronegativos (Figura 4 A, B). La proteína recombinante de aproximadamente 21 kDa obtenida del clon 1 de M. brassicae, mostró reactividad cruzada con sueros de pacientes PET/HAM y seropositivos asintomáticos para el HTLV-I pero no con los controles negativos (Figura 5). La proteína recombinante RENV se utilizó para definir mediante Western blot, la especificidad. Todos los sueros (75/75) de los individuos seropositivos mostraron una reacción cruzada con la proteína de 21 kDa extraída de lisados de M. brassicae clon 1, en contraste con aquellos de los sujetos seronegativos en los que no se observó ninguna reactividad (Figura 6). Tanto la especificidad como la sensibilidad de la prueba de ELISA para la proteína RENV fueron del 100%. No se observaron ni falsos negativos ni falsos positivos. DISCUSIÓN El principal hallazgo de este estudio fue el desarrollo de un procedimiento sencillo y altamente reproducible para la obtención de una proteína viral inmunogénica que pudiese ser utilizada como referencia en el diagnóstico de la infección por el HTLV-I. Uno de los problemas que se presenta en el diagnóstico de retrovirus humanos como el HTLV-I, radica en la sensibilidad de la prueba. Para ello, se emplean técnicas como la PCR que amplifica un molde proviral inicial, determinando el tipo de provirus HTLV-I10. Sin embargo, esta prueba de laboratorio no podría llevarse al campo para realizar estudios epidemiológicos en zonas distantes de la costa pacífica colombiana y otras regiones donde se ha informado una alta prevalencia de la infección por el virus. La elaboración de estuches diagnósticos de gran sensibilidad y especificidad, cumplen los requerimientos para diagnósticos rápidos en estas zonas; por ello se hace imprescindible la caracterización y producción de proteínas recombinantes que cumplan estas necesidades. La proteína recombinante de la gp46, producida en este trabajo, cumplió con estos requisitos. Resultados previos obtenidos en sistemas in vitro6,11 han mostrado un aumento significativo de reactividad de IgG y de IgA contra proteínas de envoltura env y gag, en pacientes PET/HAM12,13. El desarrollo de vectores de expresión que contienen secuencias de aminoácidos en la región media de la gp46 como el pB-806 (166-229 a.a.) y el pB1-806 (166-201 a.a.)5, permitieron un acercamiento en la elección de la secuencia de aminoácidos que se deseaba producir, pues presentaron niveles de reactividad significativos en sueros de pacientes infectados con HTLV-I5,14. Estos resultados demostraron que la región más inmunorreactiva de la gp62 está localizada en la porción media C-terminal de la gp46 (161- 209a.a.). El sistema de Baculovirus ha sido empleado con mucho éxito en la expresión de proteínas de envoltura del HTLV-I. En los trabajos de Arp et al.15 se emplearon estos sistemas obteniendo suficiente proteína acumulada en células de insecto. Los resultados del presente artículo junto con los estudios de inmunogenicidad de las proteínas de envoltura de HTLV-I, en especial los epítopes B y B1, indican una gran factibilidad de producir proteínas recombinantes mediante procesos rápidos, seguros, de alto rendimiento y confiables. La región media de la gp46 contiene epítopes altamente inmunogénicos. Se han comparado las reactividades relativas de anticuerpos de varias regiones de gp62 de HTLV-I usando proteínas recombinantes en ensayos de Western blot y se ha encontrado que la porción media C-terminal de gp46 es la región más inmunogénica de la proteína5. En algunos estudios se ha evaluado la reactividad de diferentes proteínas recombinantes de gp46 y se han encontrado datos importantes en el caso del epítope B1. Los pacientes PET/HAM poseen niveles altos de anticuerpos contra antígenos virales y una carga proviral alta comparada con portadores asintomáticos y pacientes ATL16. Los ensayos de ELISA y Western blot realizados en este trabajo empleando sueros de pacientes infectados con HTLV-I mostraron reactividad contra la proteína recombinante mientras RENV-21 kDa en contraste con los controles seronegativos utilizados en los que no se determinó reactividad. Con base en los resultados obtenidos en el presente trabajo, se propone el uso de la proteína RENV para el diagnóstico de la infección con HTLV-I. AGRADECIMIENTOS A las entidades financiadoras COLCIENCIAS, Fogarty Institute, Universidad del Valle y Universidad de Kagoshima. REFERENCIAS
Copyright 2002 - Revista Colombia Médica The following images related to this document are available:Photo images[rc02011f5.jpg] [rc02011f2a-c.jpg] [rc02011f4a-b.jpg] [rc02011f6.jpg] [rc02011f3.jpg] [rc02011f1.jpg] | ||||||||||||||||||||||||||
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}