 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Revista Colombia Médica, Vol. 33, No. 4, 2002, pp. 171-178 Células germinativas primordiales femeninas: origen y migración hacia los primordios gonadales Lilian Chuaire, Biol., M.Sc.¹, Magda Carolina Sánchez, Lic. Quím.²
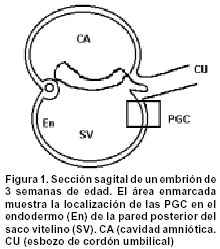
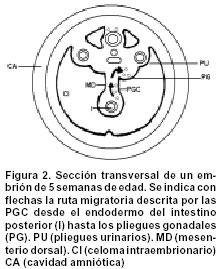
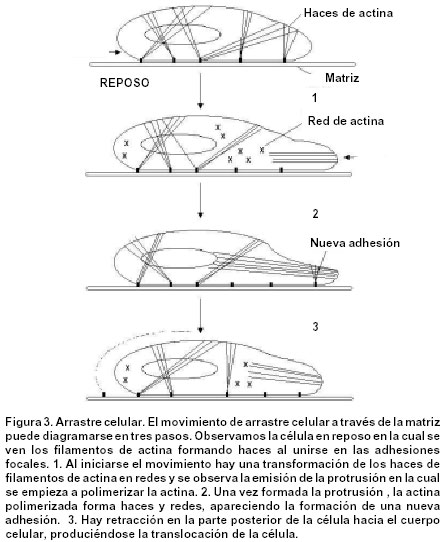
Code Number: rc02030 RESUMEN Las células germinativas primordiales femeninas (PGC), de manera semejante a como ocurre en otros mamíferos, aparecen temprano en el desarrollo, dentro de tejidos que, no sólo están alejados de las gónadas en formación, sino que, además, son de naturaleza extraembrionaria. En consecuencia, hay un proceso de translocación que las conduce hacia los primordios gonadales, donde proliferan e interactúan con células somáticas, para dar comienzo a la foliculogénesis ovárica. ¿Poseen las PGC algún tipo de “memoria” que les indique su destino? ¿Cuáles sucesos determinan la dirección de su desplazamiento desde su remoto lugar de origen, y cuál es su naturaleza? El presente artículo procura dar respuesta a estos interrogantes. Para esto se consideran temas como origen y ultraestructura de las PGC, las características de su conducta migratoria y la influencia que ejercen la matriz extracelular del mesenterio dorsal y los primordios gonadales sobre esa conducta. Palabras clave: Células germinativas. Movimiento celular. SUMMARY Female primordial germ cells (PGC), as in other mammals, appear early in the development, inside tissues that not only are far from the gonadal anlage, but also are of extra embryonic nature. Therefore, they make a translocation process that leads them to the gonad primordium, where they proliferate and interact with somatic cells in order to initiate the ovarian folliculogenesis. Do PGC possess a kind of “memory” that shows their target? What events determine the direction to which they should migrate from their place of origin? Which is the nature of these events? In this paper, we'll try to touch topics as the origin and the ultraestructure of the PGC, its migratory behaviour characteristics and the influence that the extracellular matrix of the dorsal mesentery and the developing gonad have on that behaviour. With these subjects we'll try to answer the stated questions as deep as possible. Key words: Germ cells. Cell movement. ORIGEN DE LAS PGC Las células germinativas primordiales (PGC, por sus siglas en inglés) son las células germinales que, en ambos sexos, deben emigrar y establecerse en las gónadas sexualmente indiferenciadas. Aunque en la actualidad hay buenas evidencias sobre el origen extragonadal de las PGC, durante largo tiempo hubo discusiones alrededor de dos hipótesis: Según la primera, sustentada por Waldeyer1 en 1870, en los mamíferos, las oogonias (células germinativas femeninas diferenciadas) se derivaban del epitelio superficial del ovario, llamado, por tanto, epitelio germinal; y, la segunda, propuesta por Nussbaum2 en 1880, afirmaba que las PGC eran parte de una línea celular continua que se transmitía de generación en generación y se originaba, no a partir de la envoltura epitelial ovárica, sino en lugares localizados fuera de las gónadas. Ambas hipótesis se debatieron ampliamente durante algo más de 60 años3-8, sin que se pudiera llegar a un acuerdo, porque los métodos histológicos convencionales utilizados entonces no permitían diferenciar las PGC de las células somáticas circundantes. Sin embargo, en 1948, Witschi9, mediante evidencia experimental incuestionable, determinó en mamíferos, la presencia de PGC en el endodermo del saco vitelino, sitio extragonadal desde donde emigraban hacia los pliegues gonadales primitivos, aproximadamente en la tercera semana de desarrollo. La evidencia morfológica indica entonces que, en los mamíferos, las PGC se localizan, hacia la tercera semana postfertilización, en el endodermo de la pared dorsal del saco vitelino, muy cerca del alantoides10 (Figura 1). Sin embargo, el origen real de las PGC no se ha aclarado completamente. ¿En qué momento del desarrollo estas células se separan de las líneas celulares somáticas existentes y dan origen a una línea germinal? Los experimentos efectuados con blastómeras de ratón11,12 y con cilindros celulares de huevos13, han demostrado que existe una línea celular germinal que no se segrega antes del día 6, y que, posiblemente se deriva de las células que conforman el epiblasto, es decir, de la parte de la masa celular interna del blastocisto que da origen al ectodermo embrionario. Estos hallazgos fueron corroborados por Gosden14 en 1995, quien, al microinyectar células epiblásticas de ratón en blastocistos receptores, observó que eran capaces de producir células germinativas. ULTRAESTRUCTURA DE LAS PGC Las PGC son fácilmente identificables al microscopio de luz (ML), cuando se encuentran en el endodermo del saco vitelino, pues no sólo son más grandes y más claras que las células somáticas vecinas15,16, sino que, además, son basófilas17 y exhiben actividad de la enzima fosfatasa alcalina en su citoplasma periférico18. La observación de las PGC al microscopio electrónico de transmisión (MET) y microscopio electrónico de barrido (MEB), permite apreciar su forma redondeada, y un diámetro que oscila entre 15 y 20 mm16. El núcleo, esférico, ocupa una posición excéntrica y contiene una cromatina granular muy fina y un número variable de nucléolos16,19. El retículo endoplásmico rugoso es abundante, al igual que los polirribosomas y los ribosomas libres. Cerca del núcleo se observan mitocondrias esféricas con crestas lamelares, así como un complejo de Golgi pequeño16,20. Asociada con el núcleo y con las mitocondrias, se encuentra una inclusión citoplasmática muy particular, propia de las PGC -tanto femeninas como masculinas- llamada nuage (del francés: nube). La nuage está constituida por masas electrodensas esféricas formadas por material fibroso o granular, muy semejantes a los gránulos polares ricos en ARN, característicos del plasma de las PGC en invertebrados y vertebrados no mamíferos21. El significado funcional de esta inclusión se relaciona con aumento en la actividad mitótica de las PGC y con la determinación de la línea celular germinal a partir de las células somáticas del epiblasto22. Las reservas energéticas de las PGC están representadas por depósitos de glicógeno y gotas de lípidos, necesarios para la migración hacia los pliegues gonadales15,23. También se encuentran unos pocos microfilamentos, centríolos y microtúbulos16,24, así como áreas focales de contacto estrecho con las células somáticas vecinas25. MIGRACIÓN DE LAS PGC HACIA LOS PRIMORDIOS GONADALES Migración pasiva. Aproximadamente hacia la cuarta semana de desarrollo, las PGC inician un proceso de translocación que las lleva desde el endodermo del saco vitelino, a través de la matriz extracelular del mesénquima del mesenterio dorsal, hasta su localización definitiva en los pliegues o primordios gonadales15,19. Este proceso ocurre simultáneamente con una metamorfosis en la conformación del embrión, que cambia de un aspecto inicial discoide, a una configuración tubular. La nueva forma permite que el endodermo del saco vitelino sea incorporado al intestino posterior y que, las PGC, por tanto, ocupen una posición intraembrionaria16,24 (Figura 2). Una vez dentro del embrión, las PGC abandonan el epitelio del intestino posterior a través de brechas en la lámina basal epitelial24,26, y se desplazan hacia el mesénquima subyacente. Hasta este momento (quinta semana de desarrollo), el proceso de translocación de las PGC es de tipo pasivo, como se puede comprobar por sus características ultraestructurales y por su aspecto metabólico quiescente16,26. Migración activa. Usualmente a la motilidad de las células embrionarias la acompañan modificaciones en sus características ultraestructurales. Así, el patrón cinético de una célula se puede alterar, cuando sobre ella actúan factores de crecimiento27 capaces de inducir la expresión de ciertos tipos de moléculas reguladoras de la motilidad, como las proteínas del citoesqueleto y las proteínas de adhesión28,29, que determinan la aparición de un nuevo patrón ultraestructural. Durante el proceso migratorio, no sólo intervienen factores endógenos, propios de las células, si no también la matriz extracelular, gracias a interacciones de diferente índole entre sus componentes y las células migratorias25,26,29. Una vez las PGC alcanzan el mesénquima del mesenterio dorsal, adquieren un nuevo patrón ultraestructural que las capacita para desplazarse activamente hacia los pliegues gonadales, mediante movimientos de tipo ameboide16,19,24,29. Así, pues, las PGC, al iniciar la fase activa de su trayecto migratorio, modifican sus características, pues adquieren una forma alargada23, aumentan de modo marcado la actividad de la fosfatasa alcalina24, también aumenta el número de membranas de retículo endoplásmico rugoso, la envoltura nuclear se torna irregular16, y comienzan a aparecer protrusiones de tipo lamelipodio20,24,27. Estos cambios indican que las PGC, una vez que alcanzan una posición intraembrionaria, adquieren capacidad de invasión tisular, que les permite dirigirse por sí mismas, de forma activa27, hacia los primordios gonadales23,26. Tanto las protrusiones como el borde o parte posterior (Figura 3) son estructuras de gran importancia en la cinética de las células migratorias. Las protrusiones se encuentran recubiertas por una fina capa fibrilar o glicocálix, de 30 nm de espesor, asociada con el reconocimiento de componentes macromoleculares específicos de la matriz extracelular25. En su interior hay elementos del citoesqueleto, como los microfilamentos16,24,26, que interactúan de tal manera, que dan origen a movimientos coordinados de extensión y de retracción, determinantes en el proceso de arrastre celular. El arrastre celular comprende tres pasos:
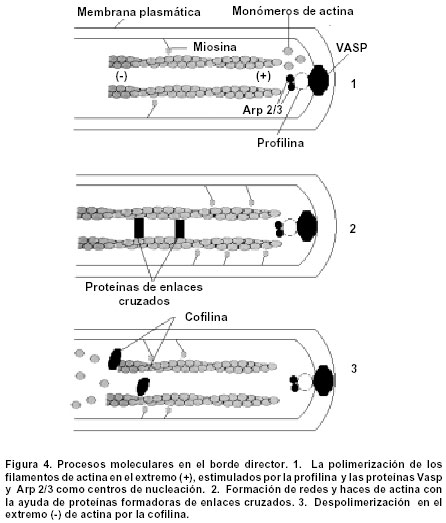
Se ha observado que la formación de las protrusiones se encuentra asociada con un proceso de polimerización de los filamentos de actina en el extremo (+) de la célula, gracias a la acción estimuladora de una proteína citosólica llamada profilina. En la dirección del ensamblaje o nucleación, también pueden participar otras proteínas, como Vasp y Arp 2/3, mientras que, en la pérdida de subunidades desde los extremos (-) de los filamentos de actina, participa la cofilina. Simultáneamente, los filamentos de actina se estabilizan como resultado de la formación de enlaces cruzados de tipo proteico, de modo que se forman retículos y haces30,32-34. Aparentemente, en el proceso se hallan comprometidas también moléculas de miosina I y II. Se ha propuesto que las moléculas de miosina I se deslizan a lo largo de los filamentos de actina en el borde director, mientras que las moléculas de miosina II se encontrarían en la parte posterior haciendo lo propio para permitir así la retracción de la célula35,36. La Figura 4 muestra un modelo hipotético que explica los eventos moleculares que tienen lugar en el borde director de las células migratorias, cuyos principios se podrían aplicar al comportamiento cinético de las PGC34,35. El resultado final de los eventos descritos, sería un ciclo coordinado con una fase inicial de extensión o alargamiento de las protrusiones, seguida por una fase de retracción, que facilitaría el desplazamiento de las PGC a través del sustrato, en este caso, la matriz extracelular35,36. Es interesante notar que, entre el borde director de las protrusiones y la parte posterior de células migratorias estudiadas, existe un gradiente de concentración de calcio, que determina el orden en el que ambas regiones permiten la interacción entre la actina y la miosina, esto, en últimas, incide en la coordinación de las fases de extensión y retracción del ciclo de motilidad celular37. Si bien la ruta migratoria descrita por las PGC desde el endodermo del saco vitelino hasta los pliegues gonadales, se ha identificado plenamente mediante métodos histoquímicos convencionales, ultracitoquímicos, inmunocitoquímicos y ultraestructurales9,24,38-42, los procesos principales a través de los que las células migratorias generan las fuerzas necesarias para desplazarse, han sido objeto de debate. Sin embargo, se acepta que el principal mecanismo generador de fuerzas para la locomoción corresponde al ensamblaje de los haces y retículos de actina en el borde director, seguido por la interacción entre la miosina y la actina tanto en dicho borde, como en la parte posterior31. Interacción entre las PGC y la matriz extracelular. Se ha determinado que la motilidad de las células embrionarias es también el resultado de interacciones locales, tanto entre dichas células, como entre ellas y la matriz extracelular vecina. Para que las PGC puedan iniciar su desplazamiento, es necesario que primero pierdan sus complejos de unión con las células somáticas del endodermo del saco vitelino, y que, además haya un sustrato idóneo que facilite la locomoción. Tal sustrato está constituido por los glicosaminoglicanos presentes en la matriz extracelular, principalmente hialuronano, condroitín-sulfato y dermatán-sulfato25,26 y por glicoproteínas estructurales o de adhesión, como la fibronectina26,42, que se caracteriza por trazar rutas migratorias celulares durante la vida embrionaria, debido a que proporciona lugares de adhesión que facilitan el avance de la célula. Es también necesario que para que las células migratorias se puedan desplazar, posean no solamente la capacidad de degradar la matriz extracelular, sino que también la puedan secretar, una vez que se establezcan en su residencia definitiva43. En la membrana plasmática de otras células migratorias estudiadas, se han encontrado proteínas receptoras de hialuronano, como CD44 u homólogas30, lo cual permite que el hialuronano recubra las células migratorias con un manto de naturaleza hidrofílica. Esta interacción es fundamental, pues confiere a las células ligadas libertad para desplazarse y para proliferar. A menudo, el cese del movimiento celular y el establecimiento de uniones intercelulares, se correlacionan con un descenso en la concentración de hialuronano presente en el tejido y con una disminución en el número de moléculas receptoras del mismo. Al mismo tiempo se ha observado un aumento en la concentración de hialuronidasa, enzima de tipo proteinasa, cuya función consiste en degradar precisamente al hialuronano43. Muchas de las enzimas de tipo proteinasa pertenecen a una de las dos clases generales: Algunas son metaloproteinasas, cuya actividad depende de la unión con el calcio o con el zinc, mientras que otras son serinaproteinasas. Ambas, metaloproteinasas y serinaproteinasas cooperan en la degradación de proteínas de la matriz, como colágena, laminina y fibronectina. Algunas de las metaloproteinasas, como las colagenasas, son muy específicas, de manera que la integridad estructural de la matriz estaría alterada por una proteólisis limitada. De esta manera, la migración celular se facilitaría por una actividad proteolítica relativamente reducida44. Existen algunos mecanismos que aseguran que la degradación de los componentes de la matriz se encuentre rigurosamente controlada:
Por otro lado, la degradación de la moléculas receptoras de hialuronano, se atribuye a la acción de enzimas de tipo metaloproteinasa, que se unen a las membranas de las células migratorias. Estas enzimas, denominadas MT1-MMP, no solamente separan de la superficie celular a la proteína receptora de hialuronano CD44 o a sus homólogas, sino que también estimulan el desplazamiento celular, pues poseen la capacidad de degradar la matriz extracelular. Las enzimas metaloproteinasas se han encontrado en células tumorales, donde promueven también fenómenos migratorios29,44,47. Influencia de los tejidos somáticos gonadales sobre la migración de PGC. Hacia la cuarta semana de desarrollo, aparecen en el embrión un par de salientes longitudinales llamados pliegues o primordios gonadales, que se sitúan a cada lado, entre el mesonefros y la raíz del mesenterio dorsal48. Al disminuir rápidamente en longitud, adquieren el aspecto de extrusiones redondeadas que se proyectan hacia el interior de la cavidad celómica26. En este momento del desarrollo las PGC comienzan a hacer su arribo a los pliegues o primordios gonadales, proceso que continúa hasta la quinta semana postfertilización (Figura 2). Los pliegues o primordios gonadales son las estructuras precursoras de la corteza ovárica. Están revestidos por un epitelio celómico en proliferación, debajo de él se encuentra un compartimiento que comprende células mesenquimáticas, vasos sanguíneos y células derivadas de los glomérulos y los túbulos mesonéfricos19,26 (Figura 2). ¿Cuál es el papel inductor que desempeñan las gónadas en formación sobre la migración de las PGC? Cuando Witschi9, demostró en 1948 el origen extragonadal de las PGC, también propuso que a estas células las guiaban y las atraían sustancias quimiotácticas producidas por los pliegues gonadales, afirmación que más tarde corroboraron Rogulska et al.49, quienes, al transplantar intestino posterior de embriones de ratón dentro de embriones de pollo, observaron que las PGC de ratón eran capaces de invadir las gónadas del receptor. Estos resultados determinaron la generación de una línea de pensamiento, pues en la actualidad, se acepta ampliamente la hipótesis de la atracción de las PGC, mediante señales específicas originadas en las gónadas en formación. Esa hipótesis se ha confirmado en especies como ratón50, donde se demostró, in vitro, que secciones de tejido gonadal podían atraer a las PGC. Igualmente, en pollos, se informó la acumulación de las PGC en regiones ectópicas, como resultado de la acción del tejido gonadal transplantado51. Por otra parte, en Drosophila, se determinó que el mesodermo gonadal expresa genes como columbus, que produce señales que atraen a las PGC52, o como wunen, comprometido en la producción de señales que repelen a las PGC, desde ciertas regiones del intestino53. Observaciones efectuadas en pez cebra54, muestran que las PGC no se desplazan directamente hacia los primordios gonadales, sino que inicialmente se dirigen hacia destinos somáticos intermedios, constituidos por células que se caracterizan por expresar el factor de transcripción wt1. Estas células somáticas producen señales que atraen a las PGC durante la somitogénesis temprana. En etapas tardías, cesa, tanto la emisión de señales, como la capacidad de respuesta por parte de las PGC, de modo que estas células migratorias pueden continuar el desplazamiento hacia su destino final. Durante el proceso de migración hacia los primordios gonadales, las PGC parecen ser guiadas por señales emitidas por células líderes que dirigen al resto de las PGC hacia su destino gonadal, como se ha observado en Drosophila55-57 y en ratones58. Mientras que en especies como el pez cebra, las PGC migran de forma individual54, en ratones, lo hacen en grupos58, debido posiblemente a que se interconectan con las células pioneras mediante extensas prolongaciones de tipo filopodial59. Aunque la migración de las PGC hacia los tejidos somáticos gonadales, depende en gran parte del mecanismo de atracción descrito, no se debe perder de vista que se trata de un proceso en el que intervienen también otros factores: Así, en Drosophila, el factor determinante en el comienzo de la migración no parece ser la atracción ejercida por el mesodermo gonadal60,61, sino, más bien la repulsión de las PGC desde regiones específicas del intestino53,55. En cuanto al mecanismo molecular que regula la migración de las PGC, se piensa que el factor de transcripción wt1, podría actuar como represor o como activador62, aunque no se ha demostrado que su acción sea indispensable en el proceso. No ocurre lo mismo con el factor steel , encontrado a todo lo largo de la ruta migratoria, cuya acción no es de naturaleza quimiotáctica, sino inductora de la motilidad, pues, unido al receptor kit de las PGC, permite la adhesión de estas células a sustratos celulares50,63. En cuanto al papel que tienen los factores de desarrollo TGF en el desplazamiento de las PGC, se ha observado que éstas son atraídas in vitro por TGF-b1.En el mismo sentido, se informó que el uso de anticuerpos anti-TGF-b1 inhibe la migración de las PGC hacia los tejidos somáticos gonadales64. CONCLUSIONES Hacia la tercera semana de desarrollo, aparecen las PGC en el endodermo de la pared posterior del saco vitelino, desde donde inician, aproximadamente hacia la cuarta semana, un trayecto migratorio que las conduce a su residencia definitiva, localizada en los pliegues o primordios gonadales. Este viaje comprende dos fases:
Otros factores para tener en cuenta, que inciden en la migración de las PGC hacia los pliegues o primordios gonadales, son las interacciones entre ellas y la matriz extracelular circundante, así como el efecto inductor ejercido por el mesodermo gonadal. La composición de la matriz extracelular, en cuanto al contenido de glicosaminoglicanos como hialuronano, que facilita el desplazamiento celular, la presencia de glicoproteínas como la fibronectina, que se caracteriza por trazar rutas migratorias, y la acción de las metaloproteinasas, que degradan no sólo componentes de la matriz como el hialuronano, sino que también separan de la membrana en las células migratorias las moléculas CD44 receptoras de hialuronano, constituyen mecanismos reguladores del movimiento celular, que pueden afectar el comportamiento cinético descrito por las PGC. La evidencia del efecto ejercido en otras especies por parte de factores quimiotácticos secretados por posibles destinos somáticos intermedios, por las gónadas en formación, y, aun por las propias PGC que actúan como pioneras en el desplazamiento, se deben tener en cuenta dentro del contexto del proceso migratorio de las PGC en el ser humano. Otros mecanismos inductores de la motilidad, como el acoplamiento del factor steel a su receptor kit, la acción del factor de transcripción wt1 y del factor de crecimiento TGF-b1, podrían también intervenir en la regulación del movimiento de las PGC hacia los pliegues o primordios gonadales. REFERENCIAS
Copyright 2002 - Revista Colombia Médica The following images related to this document are available:Photo images[rc02030f2.jpg] [rc02030f3.jpg] [rc02030f1.jpg] [rc02030f4.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}