 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Revista Colombia Médica, Vol. 34, No. 3, 2003, pp. 143-154 La genotipificación y fenotipificación de la resistencia del virus de la inmunodeficiencia humana tipo 1 (VIH-1) a los fármacos antirretrovirales Felipe García-Vallejo, Ph.D.1, Martha C. Domínguez, M.Sc.2
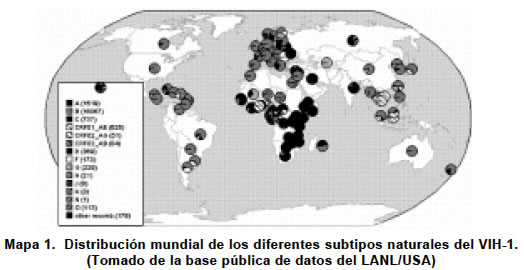
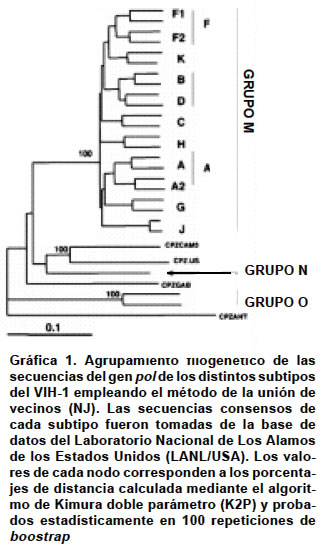
Code Number: rc03023 RESUMEN En diciembre de 2002 existía en el mundo 42 millones de personas infectadas por el virus de la inmunodeficiencia humana tipo 1 (VIH-1). Cómo se ha llegado a esta cifra, es una de las preguntas que tiene como base de respuesta la gran adaptación evolutiva del virus. Las poblaciones del VIH-1 replican como distribuciones complejas de genomas diferentes, pero genéticamente relacionadas, denominadas cuasiespecies. Así, las cepas virales que circulan alrededor del mundo presentan gran heterogeneidad de genotipos distribuidos como agrupaciones genéticas naturales con distribuciones geográficas características y dinámicas de infección y dispersión diferentes. El aumento de la capacidad de acción antiviral, facilitado por el desarrollo de nuevos fármacos, ha incrementado la frecuencia de mutantes virales resistentes. Este nuevo panorama evolutivo dificulta la aplicación de una terapia antirretroviral exitosa. Numerosos estudios retrospectivos realizados a lo largo de los años han demostrado la correlación entre la presencia de mutaciones y el fracaso terapéutico de la infección por el virus. En tal sentido ha sido importante desarrollar pruebas de genotipificación y fenotipificación que puedan discriminar las mutaciones asociadas con la resistencia a un determinado antirretroviral. Aunque no son instrumentos completamente seguros, si se convierten en herramientas poderosas para poder diseñar de manera más confiable tratamientos terapéuticos más efectivos y que retarden al máximo la aparición de cepas resistentes. Además, se pueden emplear en la vigilancia fina de la pandemia del SIDA/VIH uno de los problemas más graves de salud pública del mundo actual. Palabras clave: VIH-1. Antirretrovirales. Mutaciones resistentes. Genotipificación. Fenotipificación. SUMMARY By December of 2002 there was in the world approximately 42 millions of people infected with the human immunodeficiency virus type 1 (HIV-1). How is this number had been achieved can be partially answered on the basis on its high evolutionary capacity of adaptation. HIV-1 populations replicate as complex distribution of viral genomes genetically related termed as quasispecies. In this sense the viral strains that circulate around the world exhibit wide heterogeneity of genotypes which are distributed as natural clades with different geographical and infection dynamics. The actual progress of the antiviral capacity, in part facilitated by the developing of new antiretroviral drugs, has increased de frequency of new viral resistant mutants. This new evolutionary landscape has difficultated the approaching to a successful antiretroviral therapy. Several epidemiological studies have shown the correlation between the emergency of new mutations and the failt of antiviral therapy. In this it has been important to develop effective genotyping and phenotyping tests that screened the mutations associated with the exposition to a defined antiretroviral drug. Although these tests are not completely safe, they are powerful tools to help the design of more trustable therapies which delay at maximum the appearance of resistant mutants. Moreover genotyping tests can be used in the epidemiological surveillance of AIDS/HIV pandemic one of the most important problem of public health in the world. Key words: HIV-1. Antiretrovirals. Resistant mutations. Genotyping. Phenotyping. Los virus de la inmunodeficiencia humana (VIH) son miembros del género de los Lentivirus de la familia Retroviridae que pueden infectar tanto a humanos como a primates1. Hasta el momento se han identificado dos miembros dentro de los Lentivirus que infectan a humanos, el VIH-1 y el VIH-2. El VIH-1 es el más extendido en el mundo. De acuerdo con el informe de Barcelona de UNIAIDS se estima que a diciembre de 2002 había 42 millones de personas en el mundo infectadas por el VIH-1; aproximadamente la mitad de ellas en el Subsahara africano. En 1986, se identificó el VIH-2 a partir de 2 pacientes africanos con SIDA que no presentaban reactividad serológica frente al VIH-1, el agente clásico de la enfermedad2. La distribución del VIH-2 es más restringida geográficamente, teniendo su epicentro en África Occidental (Guinea-Bissau, Senegal, Costa de Marfil y Camerún), aunque también existe y se propaga lentamente en la India, Portugal, Angola, Mozambique y Brasil. El aumento de la capacidad de acción antiviral, facilitado por el desarrollo de nuevos fármacos, ha incrementado la frecuencia de mutantes virales resistentes. Esto desde el punto de vista terapéutico es un inconveniente, porque compromete el éxito de la terapia antirretroviral. Como una aproximación hacia un diseño más racional y personalizado de la terapia antiviral, se han estudiado los cambios mutacionales producidos por la acción de drogas antivirales sobre los genes que codifican por la retrotranscriptasa (RT) y la proteasa (P) del virus. La determinación sistemática de estas mutaciones resistentes ha ayudado al diseño de pruebas que permiten conocer el estado genético, en cuanto a resistencia antiviral, de las variantes que tiene un individuo infectado. Este conocimiento teórico a priori, permite el manejo terapéutico personalizado que asegura un mayor éxito en el tratamiento del SIDA/VIH. En esta revisión se hará una descripción de las bases genéticas de la resistencia, de los métodos para determinarla y su aplicación en el manejo integral del individuo infectado por el VIH-1. Finalmente se dará una visión personal y crítica del problema en Colombia. LA VARIACION GENÉTICA DEL VIH-1 La gran heterogeneidad genética del VIH-1 es el resultado de la elevada tasa de mutación que se genera durante la replicación del ARN viral. La tasa de mutación se define como el número de bases incorrectamente incorporadas por nucleótido y por ciclo de replicación. Representa el número de veces que la ARN polimerasa incorpora un nucleótido erróneo y es del orden de 10-3-10-5 sustituciones/nucleótido/ciclo replicativo en virus ARN3-5. La razón principal de la alta tasa de mutación es que las ARN polimerasas y las retrotranscriptasas (RT) carecen de una actividad exonucleasa 3´-5´ denominada actividad editorial4,5, así se incrementa la probabilidad de incorporación equívoca de nucleótidos cuando se compara con las ADN polimerasas que, por lo general, tienen mecanismos de corrección y reparación4. La variación genética en los virus ARN no sólo puede ocurrir por mutaciones puntuales (transiciones y transversiones), sino también por adiciones y deleciones, por recombinación homóloga y no homóloga y por reordenamiento de segmentos genómicos6. Las poblaciones de virus ARN replican como distribuciones complejas de genomas diferentes pero genéticamente relacionadas, denominadas cuasiespecies6,7. Toda población de virus ARN presenta una secuencia definida estadísticamente llamada secuencia promedio o consenso, que tiene en cada posición el nucleótido más frecuente del conjunto de moléculas de la población. Cada genoma viral puede diferir del consenso en un número distinto de posiciones nucleotídicas. Las cuasiespecies constituyen un reservorio de variantes virales, que representan un amplio rango de fenotipos con respecto a virulencia, tropismo, cinética de replicación y composición antigénica. El tamaño poblacional es de gran importancia en la heterogeneidad genética. En un individuo infectado por el VIH-1 puede existir del orden de 109 a 1012 viriones8, siendo el recambio de viriones y de células infectadas muy elevado9,10. Por tanto, el control de las enfermedades infecciosas causadas por virus ARN, entre ellas el SIDA/VIH, constituye una tarea difícil por la gran plasticidad genética y el enorme potencial evolutivo del VIH-1 y 211. Las cepas del VIH-1 que circulan alrededor del mundo presentan gran heterogeneidad de genotipos y de subtipos virales12 (Mapa 1). Los análisis filogenéticos obtenidos con base en sus secuencias génicas, principalmente de los genes pol y env, han revelado dos grandes grupos dentro del VIH-1: el grupo M (“main” o principal), subdividido en 10 subtipos hasta el momento (A-J) y el grupo O (“outlier”), con varios aislados muy divergentes entre sí12-14 (Gráfica 1). Para el VIH-2 se han descrito 6 subtipos (A-F)15,16. El subtipo A del VIH-1 y en menor medida el B, son los más frecuentes a nivel mundial. Los subtipos C, D y E, y quizá el F, podrían comportarse como no patogénicos en el ser humano. La gran expansión actual del VIH permite prever la aparición de más variantes del VIH-1 y del VIH-2 en el futuro17. La variación en la secuencia de aminoácidos de la proteína gp 120 entre los subtipos del VIH-1 es mayor que la observada en un mismo individuo o en un subtipo18,19. Dentro de un mismo subtipo, los aislados varían entre 3% y 23% en su secuencia de nucleótidos. La divergencia genética entre distintos subtipos del VIH-1 oscila en torno a 20% y 30% dentro del grupo M, y en más de 35% entre los aislados del grupo M y del O, si consideramos la secuencia genética del gen env que codifica a las proteínas de superficie19. Los virus del grupo O del VIH-1 presentan un grado de variabilidad mayor, aunque no han sido clasificados en subtipos por el bajo número de aislados caracterizados hasta la fecha20. La divergencia genética entre los subtipos del grupo M es de 14% en la región gag21. Además, las secuencias nucleotídicas del VIH-1 identificadas hasta ahora sólo tienen 50% de homología con las del VIH-212. Las distancias genéticas entre subtipos son, en promedio, dos veces mayores que para aislados separados del mismo subtipo22. Los subtipos de la mayoría de los aislados del VIH-1 son congruentes para gag y env12; sin embargo, los subtipos E y G son variantes mosaico, esto es, con env de E y G, respectivamente, pero gag de A. Todos los subtipos del VIH-1 y del VIH-2 se transmiten por vía sexual, parenteral y vertical, aunque con diferente eficiencia. De igual modo, todos pueden causar manifestaciones clínicas en las personas infectadas, aunque el VIH-2 es menos virulento, los aislados del subtipo C del VIH-1 apenas inducen formación de sincicios, una propiedad asociada con mayor virulencia; sin embargo, el curso de la enfermedad no es más favorable en estos pacientes que en los infectados por otros subtipos. RESISTENCIAS DEL VIH AL TRATAMIENTO ANTIRRETROVIRAL Desde el punto de vista genético los virus están provistos de una gran capacidad de adaptación a los cambios introducidos en su medio natural. El VIH fundamenta esta cualidad en diversos mecanismos, entre los que cabe destacar cuatro:
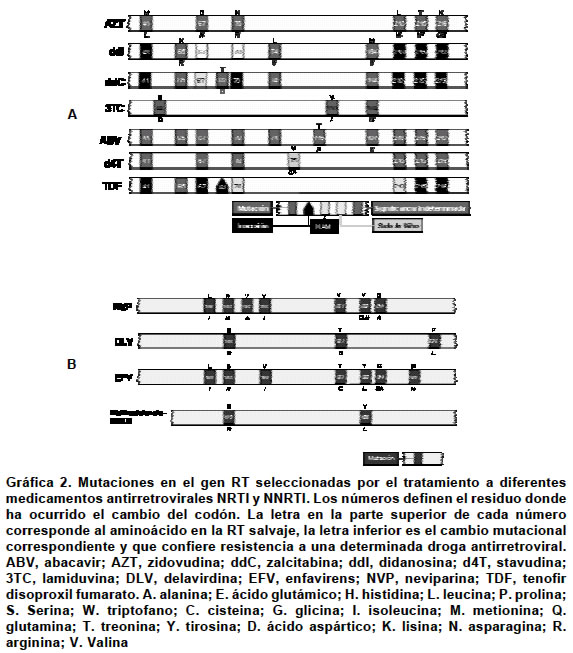
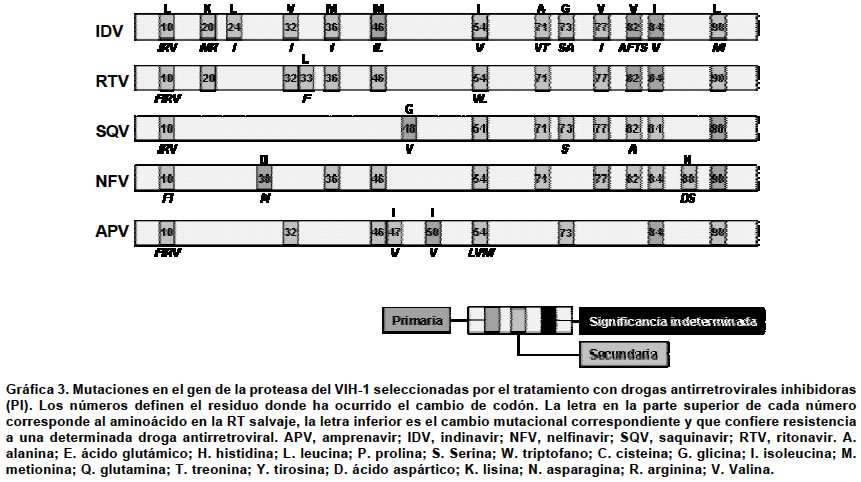
Por la notable heterogeneidad genética del VIH se producen procesos evolutivos, como la mutación que dará paso a la selección de cuasiespecies en virtud de su capacidad de supervivencia en un ambiente «hostil» (un medicamento antiviral), generándose por tanto un proceso de selección de aquellas que presenten características de supervivencia más favorables en estas circunstancias3. Como las mutaciones aparecen de forma espontánea y simplemente se seleccionan bajo la presión selectiva de los fármacos, actualmente el VIH-1 tiene capacidad de desarrollar resistencias frente a todos los antirretrovirales disponibles e incluso una capacidad potencial frente a moléculas que están por diseñarse25-27. La resistencia en sentido amplio se define como cualquier cambio que mejore la replicación del VIH en presencia de un inhibidor28. Es importante tener en cuenta que el concepto de resistencia es relativo, porque si se parte de un inóculo lo suficientemente pequeño y se usa un fármaco en concentración suficiente, un virus resistente podría presentarse como sensible. En términos específicos, la resistencia del VIH al tratamiento antirretroviral consiste en un fenotipo alterado como resultado de un cambio en el genotipo viral que se puede detectar tanto in vitro como in vivo29. Epidemiológicamente se puede hablar de resistencias primarias y secundarias11,29. Son resistencias primarias cuando se encuentran en virus de pacientes que no han sido tratados previamente, lo que implica que la infección se ha adquirido a partir de cepas de VIH-1 resistentes. Por el contrario, se trata de resistencias secundarias cuando aparecen en la población viral de un paciente como consecuencia de la presión selectiva ejercida por la exposición a fármacos antirretrovirales. TIPOS DE MUTACIONES ASOCIADAS CON LA RESISTENCIA La resistencia farmacológica al VIH-1 surge frente a la terapia antirretroviral de supresión viral incompleta y limita ampliamente el éxito a largo plazo de la terapia. Se conocen como mutaciones primarias aquellas alteraciones en el material genómico que una vez expresadas darán lugar a cambios en el sitio activo de la enzima, pues afectan la afinidad de ésta por su sustrato. Habitualmente se seleccionan pronto en el curso del tratamiento por la presión selectiva ejercida por el fármaco, en un intento de evadir su acción inhibidora del medicamento30. Las mutaciones secundarias se van acumulando en el genoma viral que ya posee mutaciones primarias con la finalidad de restaurar la ventaja cinética que ha pagado la enzima por la mutación primaria. Son seleccionadas por la ventaja replicativa que confieren y la mejora de la afinidad por el sustrato natural de la enzima. Por sí mismas, estas mutaciones secundarias poseen un efecto mínimo o nulo en la magnitud de la resistencia al tratamiento antirretroviral30. Tanto la transcriptasa reversa como la proteasa del VIH-1 han sido seleccionadas como potenciales blancos terapéuticos para su inhibición mediante fármacos. Actualmente existen una serie de medicamentos inhibidores que se han clasificado como nucleótidos/nucleósidos análogos de la RT (NRTI), no nucleósidos inhibidores de la RT (NNTRI) y los inhibidores de la proteasa (PI). La transcriptasa reversa retroviral es una ADN polimerasa dependiente de moldes de ARN que cataliza la síntesis del ADN proviral. La enzima es un heterodímero conformado por las subunidades p66 y p51. La subunidad p51 posee 440 residuos de aminoácidos mientras que la p66 tiene 560 residuos. Aunque p51 y p66 comparten 440 residuos sus plegamientos tridimensionales son diferentes. La subunidad p66 contiene los sitios de unión a la curvatura del ADN sustrato y el sitio activo que cataliza la formación de enlaces fosfodiester entre nucleótidos 5´-trifosfato (dNTP) libres y el extremo 5´ de la cadena de ADN que está siendo sintetizada; por el contrario la p51 no tiene ninguna actividad catalítica y sirve como andamio para la actividad enzimática de la p66. La subunidad p66 tiene cinco subdominios que incluyen los de los dedos, la palma y la muñeca que participan activamente en la reacción de polimerización, además de los subdominios conector y de ARNsa H. La mayoría de las mutaciones resistentes se localizan en la región de la polimerasa 5´ y en particular en los dedos y en la palma31,32. En la Gráfica 2a y b se presentan las principales mutaciones que cambian codones de la secuencia de aminoácidos de la RT salvaje y que exhiben resistencia a los NRTI (zidovudine, didanosine, zalcitibine, stavudine, lamivudine, y abacavir) y a los NNRTI (nevirapine, delavirdine y enfavirenz). El tenofir disoproxil fumarato (Tenofir DF) es el único nucleótido análogo inhibidor de la RT que ha sido aprobado por la FDA. Recientemente se han presentado resultados de los ensayos clínicos de este inhibidor y de un análisis integrado de resistencia genotípica33,34. Los estudios 902 y 907 eran de carácter aleatorizado y controlados con placebo en pacientes con carga viral mayor de 400 copias/ml que recibieron terapia antirretroviral estable más de 8 semanas y luego añadieron a su régimen tenofovir DF o placebo. Después de 24 semanas, se cambió de grupo a los pacientes asignados a placebo para que recibieran tenofovir DF. Hasta el momento se dispone de datos a la semana 48 de terapia con tenofovir DF de 333 pacientes que inicialmente recibieron el fármaco y de 103 que cambiaron desde placebo a tenofovir DF. Se determinó una resistencia extendida a nivel basal, pues casi 75% de los pacientes tenía MAT. En conjunto, la reducción de la carga viral media después de 48 semanas de terapia fue aproximadamente de 0.6 log10 copias/ml (una reducción de 4 veces). Los pacientes con la mutación L210W presentaban un índice de respuesta marcadamente inferior y una disminución de la susceptibilidad fenotípica. La L210W fue casi siempre encontrada en asociación con la T215Y (en más de 98% de los casos) o la M41L (en más de 94%); por ello, la presencia de la L210W era diagnóstico de 3 ó más MAT, incluyendo la M41L y la L210W. Los índices de respuesta en pacientes con un patrón alternativo de MAT (la D67N, la K70R y las K219Q/E/N) funcionaron mucho mejor, con sólo cambios menores en la susceptibilidad fenotípica. La mutación M184V asociada con la resistencia a la lamivudina se relacionó con un aumento de la respuesta virológica35. Está pendiente por determinar qué importancia tienen para tenofovir otros patrones de mutaciones de ITIN observados con frecuencia junto con otras mutaciones identificadas recientemente como importantes para la respuesta de los ITIN como la 69D, la 44D y la 118I36,37. La proteasa del VIH-1 es la enzima responsable por el procesamiento postraduccional de la poliproteínas precursoras virales gag y gag-pol que generan las proteínas estructurales del virión y las enzimas retrovirales respectivamente. La enzima es una aspártico proteasa compuesta por dos monómeros idénticos no covalentemente asociadas con 99 residuos38. Su sitio activo contiene una tríada catalítica conformada por Asp-Thr-Gly en las posiciones 25 a 27. En la actualidad existen seis inhibidores de la proteasa que aprobados por la FDA: amprenavir, indinavir, lopinavir (manufacturado en combinación con ritonavir), nelfinavir, ritonavir y saquinavir. El espectro de mutaciones seleccionadas por el tratamiento con estos inhibidores sobre la proteasa está bien caracterizado y se resume en la Gráfica 3. JUSTIFICACIÓN DE LA DETERMINACIÓN DE RESISTENCIAS DEL VIH Para el tratamiento de la infección por el VIH-1 se han aprobado 16 medicamentos antirretrovirales: 7 son análogos de nucleótidos/nucleósidos que son inhibidores de la RT (NRTI); 6 inhibidores de la proteasa (PI) y 3 son no nucleósidos inhibidores de la RT (NNRTI). En individuos que no han sido tratados previamente, el empleo de dos o más combinaciones de clases de inhibidores, terapia de alto rendimiento (HAART), produce una prolongada supresión y una reconstitución inmunológica. Numerosos estudios retrospectivos realizados a lo largo de los años han demostrado la correlación entre la presencia de mutaciones y el fracaso terapéutico37,38. Recientemente se han llevado a cabo estudios prospectivos que comparan la evolución de pacientes distribuidos en dos cohortes: en un grupo pacientes tratados según recomendaciones empíricas generales y en el otro pacientes cuyo tratamiento es guiado por los resultados de las pruebas de resistencia al tratamiento antirretroviral. Los resultados de este último tipo de estudios avalan la realización de dichas pruebas para la selección del tratamiento más eficaz en los pacientes previamente tratados en los cuales no se han obtenido respuesta virológica completa39. En el Cuadro 1 se muestran los principales ensayos aleatorizados controlados que han demostrado el impacto a corto plazo de la detección de resistencia a los antirretrovirales38. PRUEBAS PARA LA DETERMINACIÓN DE RESISTENCIAS Existen dos grandes grupos de técnicas para la determinación de resistencias del VIH al tratamiento antirretroviral: las genotípicas y las fenotípicas (Cuadro 2). Las pruebas genotípicas tienen como base el análisis del genoma y por tanto encuentran la presencia de mutaciones. En función del principio en que se basen, el número de mutaciones detectables es distinto. Las técnicas que utilizan la secuenciación detectan todas las mutaciones presentes en las regiones del genoma del VIH que codifican para la retrotranscriptasa y para la proteasa. Cuando el fundamento de la técnica es la hibridación, sólo se encuentra determinado número de mutaciones de significación conocida40. Las pruebas fenotípicas consisten en sistemas de replicación in vitro que enfrentan al virus con diferentes concentraciones de fármacos antirretrovirales (Cuadro 2). El grado de inhibición del crecimiento se establece por comparación con una cepa de referencia41. PRUEBAS GENOTÍPICAS COMERCIALES Existen tres pruebas genotípicas comerciales, la secuenciación directa de ARN retroviral y dos técnicas basadas en la hibridación (LiPATM y GeneChipTM), cuyo fundamento se describe brevemente a continuación. Secuenciación. El principio general de esta técnica consiste en la obtención de moléculas de ADN un nucleótido más largo que la precedente y su posterior separación mediante electroforesis en gel de poliacrilamida (PAGE) de alta resolución42. En la actualidad, aunque el principio básico no ha cambiado, las técnicas modernas de secuenciación se derivan del método de Sanger43, según el cual en lugar de fraccionar el ADN en cuestión, se procede a la síntesis de pequeños fragmentos a partir de la cadena molde. Esta síntesis tiene una peculiaridad, que es la incorporación de nucleótidos terminadores de cadena. Estos nucleótidos en realidad son didesoxinucleósidos-5´-trifosfato (ddNTP), es decir, carecen de grupo hidroxilo (OH) en posición 3', que es fundamental para la elongación de la cadena, porque es precisamente en esta posición donde la polimerasa inserta el siguiente nucleótido durante la síntesis de ADN. La adición de la proporción adecuada de nucleótidos naturales con respecto a estos didesoxinucleótidos dará lugar a la síntesis de fragmentos de diversos tamaños. Para secuenciar el VIH se pueden adoptar dos estrategias: la secuenciación del genoma proviral integrado en los linfocitos, que ya está como ADN, o la realización de una retrotranscripción previa de ARN viral plasmático, para obtener una colección de cADN retrovirales. Para poder obtener secuencias confiables, un requisito fundamental es la idoneidad del material de partida, tanto en pureza como en cantidad. Para garantizar este aspecto se realiza, en primera instancia, una PCR «anidada», que no es otra cosa que dos PCR seguidas, de forma que el amplificado conseguido es mucho más abundante que si se realizase una PCR ordinaria. Tras la obtención del material genético suficiente se procede a la purificación del amplificado para eliminar restos de iniciadores, sales, enzimas, nucleótidos, etc. Una vez obtenidos todos los fragmentos se procede a su separación electroforética, que se realiza de forma automatizada en un secuenciador; ésta puede ser según la clásica PAGE de alta resolución o mediante electroforesis capilar. Independientemente de la técnica de electroforesis acoplada al secuenciador, los fragmentos se separarán en función de su tamaño molecular. Las señales se recogen en un procesador electrónico acoplado al secuenciador y éste deduce la secuencia por comparación de los fragmentos solapantes, de manera que esta secuencia en realidad es una secuencia consenso que representa las secuencias de las subpoblaciones virales mayoritarias. Para determinar la presencia de mutaciones que confieren resistencia a los antirretrovirales se extrae el genoma de una muestra del paciente y se procede a su secuenciación. Una vez obtenida la secuencia del VIH de la muestra problema, la secuencia obtenida del gen RT o de la proteasa se compara con las correspondientes secuencias de una cepa salvaje del VIH prototipo no resistente utilizando un programa informático acoplado al secuenciador44,45. LiPATM. El fundamento de esta técnica es una hibridación reversa post-PCR45-47. El soporte de la hibridación consiste en tiras de nitrocelulosa con sondas de oligonucleótidos específicos inmovilizadas en líneas paralelas. Estas sondas son complementarias a la secuencia de mutaciones conocidas que confieren resistencia a los fármacos antirretrovirales. La amplificación de la muestra se realiza con oligonucleótidos cebadores biotinilados que permitirán el revelado de la reacción. Después de la hibridización se añade un conjugado compuesto por estreptavidina-fosfatasa alcalina, que se unirá a cualquier híbrido formado sobre la tira. Al añadir el sustrato de la enzima se producirá un precipitado marrón-púrpura en las posiciones correspondientes. VIH GeneChipTM (Affimetrix). A pesar de que el fundamento de esta técnica es una hibridación, el resultado final es la secuenciación del material amplificado a partir de la muestra. El soporte de hibridación son microarreglos de ADN o «chips» de silicio que contienen un «biblioteca combinatoria» de sondas, que consiste en una gran cantidad de oligonucleótidos solapantes (409.000/1.28 cm2). La hibridación de estos microarreglos con cADN retroviral va a permitir identificar el nucleótido presente en cada posición de la secuencia a analizar48,49. Como los productos de RT-PCR están marcados, permitirá deducir la secuencia la detección de fluorescencia tras la hibridación mediante barrido de láser e integración de las señales obtenidas con un programa informático. Una ventaja de esta técnica es que permite el análisis simultáneo de un elevado número de muestras. Pruebas fenotípicas comerciales. Como las pruebas genotípicas son una medida indirecta, pues la presencia de mutaciones no siempre implica su expresión, las pruebas fenotípicas deberían ser el primer escalón natural en la detección de resistencias50,51. Básicamente estas técnicas consistían en el cocultivo de linfocitos del paciente con líneas celulares linfoides, como MT2 y MT4, en presencia de fármacos antirretrovirales52. Transcurridas unas semanas se medía la replicación viral por la producción de antígeno p24 ó la actividad retrotranscriptasa. Estas técnicas tienen varias limitaciones e inconvenientes como son la laboriosidad, altos costos, la infraestructura necesaria, el largo tiempo requerido y la falta de estandarización. Todo esto ha sido resuelto en parte por las técnicas de virus recombinantes (RVA)53. Las técnicas de virus recombinantes determinan la concentración que inhibe 50% de la infectividad (IC50) y la comparan con la IC50 de una cepa de referencia o de un aislamiento anterior de un mismo paciente. Actualmente, existen dos técnicas comerciales que se diferencian sobre todo en los siguientes aspectos:
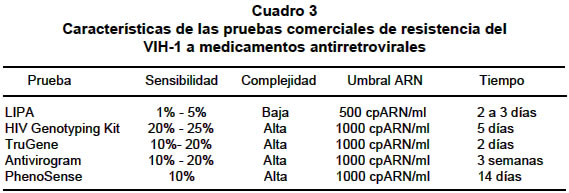
Entre las principales ventajas que presentan las pruebas fenotípicas de resistencia a los fármacos antirretrovirales están su capacidad para medir directamente la sensibilidad al fármaco en cuestión, de forma importante aportan información sobre resistencias cruzadas, que se pueden aplicar a cualquier molécula y que son relativamente fáciles de interpretar. Limitaciones de las pruebas de detección de resistencias. Una precaución importante para tener en cuenta a la hora de tomar las muestras para determinar resistencia, es asegurar que en ese momento el paciente continúa con el tratamiento que supuestamente ha fracasado. Algunos autores54 recomiendan que el plazo máximo transcurrido entre el cese del tratamiento y la detección de resistencias no supere los 15 días, pues el rápido recambio de la población viral conduciría a un predominio de subpoblaciones salvajes que daría lugar a falsos negativos en la determinación de resistencias55. Dentro de las limitaciones que en general presentan estas técnicas, independientemente del principio en que se basen, está la proporción mínima que debe representar una subpoblación determinada con respecto al global de la muestra56. A pesar de que varía en función de la sensibilidad de la técnica, se puede decir que se necesita una proporción de 20% para que esa subpoblación esté representada en el conjunto y por tanto sea detectable. Además, se requiere un umbral de carga viral mínimo para garantizar la obtención de resultados fiables, que por lo general se establece en 1.000 copias de ARN/ml, aunque algunas técnicas aseguran que se podría rebajar este umbral a 500 copias de ARN/ml (Cuadro 3). Como ya se ha dicho, ambos tipos de técnicas de determinación de resistencias poseen ventajas y desventajas, pero en principio las genotípicas parecen más rápidas, asequibles y baratas, aunque utilizar sus resultados para predecir la respuesta a los fármacos puede resultar un poco prematuro por varios factores: presencia de mezclas de subpoblaciones no determinadas, conocimiento incompleto de las mutaciones que confieren resistencia, interacción de diferentes mutaciones, resistencias cruzadas y respuesta variable a los fármacos en función de los pacientes. Por eso, es necesario definir mejor el valor predictivo de este tipo de técnicas. Por el contrario, las fenotípicas son una medida directa de la resistencia, pero su interpretación también es difícil. Poseen la ventaja de determinar la influencia de mutaciones múltiples en el genoma, a diferencia de las técnicas genotípicas, pero también por ello puede quedar enmascarado el papel individual de una mutación determinada. Otro inconveniente es que para llegar a determinar fenotípicamente una resistencia es necesario que una proporción suficientemente grande de la muestra posea algunas mutaciones (Cuadro 3). RECOMENDACIONES PARA EL MANEJO DE LAS PRUEBAS DE RESISTENCIA Desde diferentes organizaciones internacionales se han emitido recomendaciones para la detección de resistencias a los antirretrovirales. Entre las guías internacionales se destacan cuatro como ineludibles: las emitidas por el Department of Health and Human Services (DHHS) de los Estados Unidos57, las de la British HIV Association58, las publicadas por Hirsch et al.59 y por Schinazi et al.60 La incorporación selectiva de las técnicas genotípicas a la práctica asistencial tiene interés, primero, por el beneficio individual de cada paciente, pues su filosofía es optimizar e individualizar el tratamiento. En sentido colectivo, esta incorporación también tiene utilidad pues permite conocer la «epidemiología» de las cepas resistentes del VIH, incidir en la mejor utilización del arsenal terapéutico disponible e intentar disminuir el riesgo de aparición de resistencias potencialmente transmisibles. Aspectos pendientes. Desde el punto de vista de los presentes autores existen al menos seis aspectos relativos con la determinación de resistencias del VIH cuyo esclarecimiento debe ser prioritario (Cuadro 4). Es preciso establecer cuáles son los beneficios constatables a largo plazo en términos de evolución virológica. A continuación se delimitará en qué situaciones debe optarse por una prueba genotípica o fenotípica. En tercer lugar, convendría describir las características intrínsecas de sensibilidad de las diversas técnicas para detectar subpoblaciones virales minoritarias. Es también oportuno disponer de un método de evaluación específico para delimitar las resistencias atribuibles a cada fármaco en el contexto de regímenes combinados. Otro punto pendiente de resolución es establecer la cadencia de las determinaciones de resistencias y las resistencias cruzadas. En última instancia, parece inexcusable impulsar estudios de costo-efectividad en la aplicación asistencial de las pruebas de resistencia. Uno de los aspectos relativos con la determinación de resistencias que por su trascendencia requiere un tratamiento especial, es la interpretación de los resultados de estas técnicas61. No existe una pauta acerca de cuál sea la mejor interpretación de las pruebas. Entre las posibilidades disponibles se encuentra la interdisciplinaria, que implica la participación de microbiólogos, clínicos y farmacólogos62 y los sistemas basados en reglas generadas por la evidencia científica y la realización del denominado «fenotipo virtual» (Virco™), que es una algoritmo de búsqueda por confrontación de las mutaciones generadas por un tratamiento en una gran base de datos que correlaciona una determinada mutación con un fenotipo resistente específico. En la práctica se usan estos tres tipos de interpretaciones, que probablemente aportan sus beneficios, pero se necesitan estudios que las comparen con el fin de hallar el mejor método. Tanto las pruebas genotípicas como las fenotípicas presentan problemas a la hora de interpretar sus resultados. Las pruebas genotípicas aportan información sobre «posibles» resistencias, en el sentido de que no se puede garantizar la expresión de las mutaciones determinadas y por ello requieren guías de interpretación basadas en la mejor evidencia científica disponible63. Además, estas técnicas poseen mayor valor para descubrir resistencias, considerando las mutaciones presentes, que para predecir sensibilidad sería necesario descartar todas las posibles causas de resistencia. Un hecho que reviste especial dificultad es la interpretación de patrones de varias mutaciones, multirresistencia64. Por su parte, las técnicas fenotípicas constituyen una medida de la replicación viral en presencia de fármacos antirretrovirales y ésta responde a diversos factores, unos conocidos y otros no60. Además, en este sentido, los puntos de corte para determinar la resistencia no están ni bien definidos ni clínicamente validados. Otro factor que dificulta la interpretación de los resultados es el desconocimiento de los niveles mínimos requeridos in vivo para lograr la supresión de la replicación viral65, que resulta aún más complejo en el caso de los inhibidores de la retrotranscriptasa análogos de nucleósido, pues estos se activan por trifosforilación en la célula blanco, de manera que la concentración activa de fármaco ha de medirse en el interior de la célula, con la dificultad que esto entraña. Se debe considerar el hecho de que existen distintas causas del fracaso del tratamiento antirretroviral: el paciente, el fármaco y el VIH. La principal causa de fracaso atribuible al paciente es el mal cumplimiento de la pauta prescrita. De los aspectos relativos al fármaco se destaca, tanto parámetros farmacocinéticos como otros factores farmacológicos. Dentro de los primeros, el más directamente asociado al fracaso es una absorción insuficiente del fármaco, que condicionará concentraciones plasmáticas subterapéuticas. Con respecto a los factores farmacológicos, se encuentran circunstancias asociadas con el fracaso virológico como una insuficiente potencia antiviral intrínseca de la combinación de fármacos, interacciones de antirretrovirales y otros fármacos, y una activación farmacológica inadecuada. Por último, el fracaso puede estar condicionado por causas atribuibles al propio VIH-1, como son la replicación en compartimientos «santuario» y el desarrollo de resistencias38. Por tanto, a pesar de la importancia que posee la determinación de resistencias al tratamiento antirretroviral, no se debe olvidar que existen otras causas de fracaso que se deben también explorar. Este campo está en constante evolución, que exige una continua puesta al día debido al desarrollo de nuevos fármacos y nuevos blancos terapéuticos, la descripción de nuevas mutaciones asociadas con la resistencia y la acumulación y jerarquización de evidencia científica. Existen bases de datos de alimentación continua y fácil consulta a través de varias sitios del Internet, de mutaciones que confieren resistencia al VIH que en este sentido facilita enormemente la actualización de los conocimientos. Asimismo, las técnicas de determinación de resistencia se verán obligadas a asumir este dinamismo, adaptándose a los datos que se vayan generando. Por tanto, cada vez se debe tener en cuenta más factores a la hora de interpretar los resultados de la determinación de resistencias del VIH a los fármacos antirretrovirales. CONSIDERACIONES FINALES La pandemia del SIDA/VIH se perfila como uno de los problemas más agudos de la salud pública mundial de este milenio. La dinámica mutacional del VIH-1 asociada con factores ambientales naturales o impuestos, hace casi imposible en las condiciones actuales, la cura de esta enfermedad. Un aspecto preocupante de esta situación es la gran amplificación en la transmisión de cepas cada vez con un mayor espectro de resistencia, multirresistencia, que obliga al desarrollo de nuevos esquemas terapéuticos. Infortunadamente los países del tercer mundo en los que la prevalencia del virus es más alta que en la mayoría de los países desarrollados, se debaten entre la pertinencia del tratamiento y el costo de la terapia antiviral. Es importante destacar que en los países tercer mundistas, la aplicación de monoterapia o de combinaciones que se sabe desarrollaran rápidamente resistencia a las terapias, es la que el Estado y los sistemas de medicina asistencial ofrecen casi a regañadientes. Tal como se ha demostrado, este tipo de terapia puede ser una solución parcial al problema, sólo si se complementa con el seguimiento genético de la resistencia asociada. Infortunadamente los elevados costos de estos métodos hacen casi imposible su aplicación por los sistemas de cubrimiento de salud. Epidemiológicamente en Colombia no existen estudios completos que muestren cuál es la circulación de los subtipos virales, ni mucho menos de las variantes resistentes; esto produce una absoluta falta de conocimiento para el manejo de este gran problema de salud pública nacional. Además existen muy pocas opciones para brindarle al portador del VIH-1 un manejo terapéutico integral adecuado. La monoterapia es cosa del pasado; su ineficiencia no compensa los «menores costos» ante lo que ofrece la terapia múltiple y de alto rendimiento TAR. En este sentido el paciente está en una desventaja clínica pues el éxito terapéutico de la terapia que se administra en el sistema público de salud y en las EPS, está obsoleta y aboca al paciente a una disminución progresiva y acelerada de su condición física contraviniendo el derecho fundamental a una vida normal. REFERENCIAS
Copyright 2003 - Revista Colombia Médica The following images related to this document are available:Photo images[rc03023g3.jpg] [rc03023t3.jpg] [rc03023g2.jpg] [rc03023t4.jpg] [rc03023m1.jpg] [rc03023t2.jpg] [rc03023g1.jpg] [rc03023t1.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}