 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Revista Colombia Médica, Vol. 35, No. 2, 2004, pp. 101-111 Clonación animal: avances y perspectivas Lilian Chuaire, M.Sc.1, Magda Carolina Sánchez, Lic. Quim.2, María Liliana Franco3
Recibido para publicación enero 8, 2004
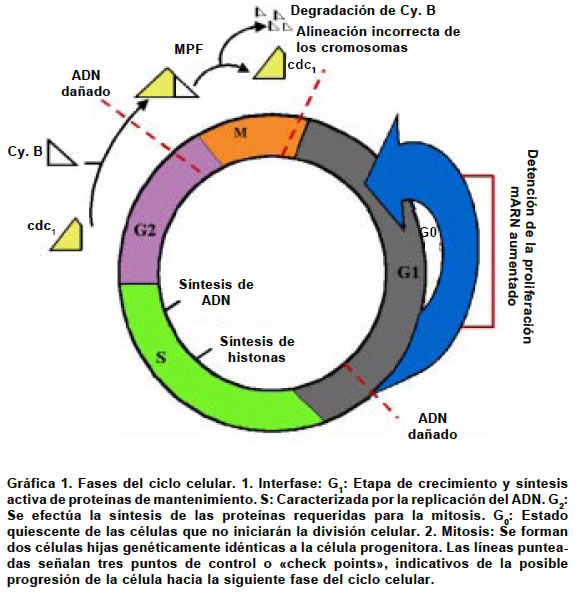
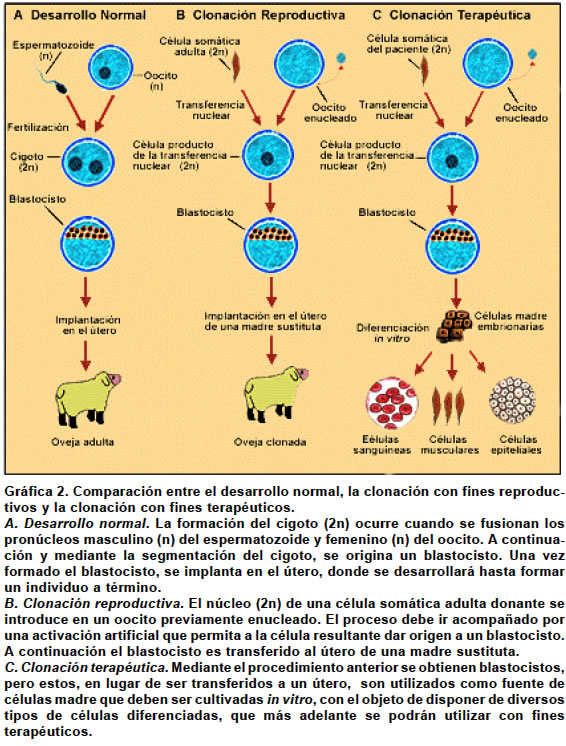
Code Number: rc04017 RESUMEN Pocos avances han revolucionado tanto la biología del desarrollo como la clonación animal. A partir del nacimiento de la oveja Dolly en 1996, primer clon derivado de un animal adulto, se inició una nueva era científica caracterizada por una creciente desmitificación de las células diferenciadas como entes inmutables en su organización nuclear y estructura cromatínica, y por una mejor comprensión de los mecanismos que regulan el desarrollo. Este trabajo revisará algunos de los logros y de las limitaciones de las técnicas utilizadas, tanto en la clonación terapéutica como en la reproductiva, así como las perspectivas que su aplicación permite vislumbrar hacia un futuro próximo. Igualmente se reseñan algunas consideraciones que han sazonado el debate ético sobre este controversial tema. Palabras clave: Clonación de organismos. Clonación embrionaria. Clonación humana. Ingeniería genética. Ética. SUMMARY Few recent advances have revolutionized the developmental biology as the animal cloning has. Since the birth of Dolly, the sheep, in 1996, which was the first derived clone of a mature animal, a new scientific era began. It has been characterized by growing demystification that differentiated cells are unalterable entities in its nuclear organization and chromatin structure, and by a better understanding of the mechanisms that regulate the development. Throughout this paper, we will review some of the achievements and limitations of the techniques used, both in therapeutic and in the reproductive cloning, as well as the perspectives that its application allows to glimpse within a close future. At the same time, we will point out some considerations regarding the ethical debate that surrounds such a controversial issue. Key words: Cloning organisms. Embryo cloning. Human cloning. Genetic engineering. Ethics. Los seres humanos son producto de un proceso evolutivo que comenzó hace aproximadamente 3,900 millones de años1-4, cuando aparecieron sobre la faz de la tierra las primeras criaturas unicelulares capaces de vivir sin oxígeno y adaptadas a las altas temperaturas reinantes. Según Domínguez et al.5 “la evolución es el proceso por el cual los seres vivos se han originado los unos de los otros por descendencia y cambios más o menos graduales a lo largo del tiempo y originándose por transformación especies nuevas”. El producto más perfecto de este proceso, siempre en marcha, es el hombre. Los antecesores del ser humano se deben buscar en las primeras formas de vida que aparecieron sobre la tierra, esto es, en microorganismos muy simples, que, con el transcurso de tiempo, adquirieron una mayor complejidad, en la medida en que experimentaron transformaciones que les permitieron interactuar más favorablemente con el entorno. ¿Cómo ocurrió la evolución de los seres vivos? Este fue un misterio sin resolver durante siglos. Sin embargo, la inteligencia del hombre desarrolló una herramienta útil para la comprensión, tanto de la naturaleza del universo y de todos los seres vivos que lo habitan, como del problema central de la evolución: esa herramienta es la ciencia, en otras épocas llamada “filosofía natural”6. A través de la ciencia, el hombre logra avanzar en el descubrimiento de la realidad de los seres vivos y en la resolución del problema evolutivo. La célula es uno de los hallazgos más significativos que el hombre ha efectuado a lo largo de su constante búsqueda. Cada célula es un universo que contiene la información necesaria para que se manifiesten las características físicas de un individuo. La información reside en los genes, estructuras que son objeto de estudio de la disciplina científica conocida como Genética. De la mano de la ciencia genética va la tecnología, por cuenta de la ingeniería genética, que se ocupa de la transferencia de genes de un organismo a otro y de crear nuevos genes para nuevos individuos potenciales creados a voluntad. También está la Genómica, que proporciona los datos necesarios para la ingeniería genética, como los genes que se han de transferir, dónde hallarlos y cómo actúan. Por otro lado aparece la clonación, tecnología que permite la aplicación de todo el poder de la ingeniería genética y de la genómica en animales y en plantas6. ¿QUÉ SIGNIFICA "CLONACIÓN"? La palabra clonación (del griego klon: retoño) tiene diferentes significados. En su acepción más común, significa la obtención de uno o de varios individuos, bien sea a partir de una célula (diferenciada o indiferenciada), o simplemente, a partir de un núcleo. Los individuos así clonados son idénticos o casi idénticos al original7. En un sentido más estricto, dentro del contexto de la ingeniería genética, la clonación consiste en aislar y amplificar o multiplicar un gen determinado o de un segmento de ADN, procedimiento que se lleva a cabo dentro de un tubo de ensayo. CLONACIÓN ANIMAL En la naturaleza, hay dos formas de reproducción celular: sexual y asexual. En la reproducción sexual, propia de los organismos pluricelulares, ambas células germinativas, la femenina u oocito y la masculina o espermatozoide, se fusionan para formar una nueva célula, llamada cigoto. Esta nueva célula no sólo comienza a diferenciarse, sino también se multiplica, mediante el mecanismo de reproducción asexual llamado mitosis. Un individuo adulto será el resultado de las sucesivas divisiones mitóticas experimentadas por el cigoto. La reproducción de tipo sexual confiere una ventaja competitiva a los organismos que la poseen, pues al ocurrir un proceso de recombinación génica de carácter aleatorio, aumenta la probabilidad de la descendencia para sobrevivir en ambientes de variabilidad imprevisible8. Los organismos pluricelulares son pues, clones ordenados de células con un mismo genoma, pero especializadas hacia funciones distintas. Esto indica que cuando se forma el cigoto, su repertorio de actividades, aunque limitado, es suficiente para dar origen a organismos con una elevada complejidad estructural. El grado de complejidad dependerá de lo que se conoce como proceso de diferenciación. Se entiende por diferenciación, el resultado de la expresión o de la inhibición de determinados grupos de proteínas, con la consecuente formación de tejidos. Los tejidos son grupos de células somáticas especializadas, que debido al proceso de diferenciación experimentado, están inhabilitadas para dar origen a nuevos individuos9. Es claro que el asunto de fondo en la clonación tiene sus raíces en las primeras etapas del proceso evolutivo, cuando los microorganismos existentes comenzaron a producir, mediante mitosis, copias exactas de sí mismos. Así, una célula somática (no sexual), se podía replicar a sí misma, de modo que el material genético de las células hijas resultaba idéntico al de la célula progenitora. En ese orden de ideas, se podría considerar que la mitosis es un tipo de clonación “natural” que no necesita manipulación alguna. CLONACIÓN ARTIFICIAL En contraste con la llamada clonación natural, las técnicas seguidas para la clonación celular artificial, requieren un proceso de elaboración o de manipulación que permite obtener copias idénticas o casi idénticas de las células madre o progenitoras utilizadas. Si el producto o embrión se transfiere a un útero, se produce la implantación en el endometrio y se desarrolla un nuevo ser (clonación reproductiva). Pero si se transfiere a un medio de cultivo, el embrión dará origen a células madre embrionarias con la potencialidad para diferenciarse hacia cualquier tipo de célula adulta (clonación terapéutica). MÉTODOS DE CLONACIÓN 1. Partición. En esta técnica se utilizan embriones octocelulares, en estado de preimplantación. A partir del embrión seleccionado, se toman mitades o secciones que posteriormente se introducen dentro de zonas pelúcidas naturales o artificiales. A continuación, se efectúa la implantación del producto en el endometrio. El número máximo de células del embrión, no puede ser superior a 8, porque a partir de este momento, se inicia la expresión del genoma embrionario. Los individuos obtenidos son prácticamente idénticos entre sí, aunque diferentes a los progenitores, por lo cual se considera que son el equivalente de los gemelos monocigóticos10. Esta técnica se ha seguido ampliamente para la clonación de animales de granja11. Como ejemplos de esta técnica están las ovejas Megan y Morag, del Roslin Institute6. 2. Clonación por transferencia nuclear de células somáticas (SCNT: Somatic Cell Nuclear Transfer). Los requisitos mínimos para la SCNT incluyen el uso de dos tipos de células: somáticas o no sexuales, y sexuales femeninas, u oocitos. Los núcleos de células somáticas de individuos postnatales se transfieren dentro de oocitos o de cigotos enucleados12-14. Esta técnica tiene la ventaja que permite conservar el genoma durante la diferenciación celular y la capacidad del citoplasma celular para reprogramar la actividad génica y aumentar la redireccionalidad de la diferenciación celular. Sin la aplicación de estos dos principios, la clonación no es posible15. Para clonar a un ser vivo mediante SCNT, se extrae el material genético de ambos tipos de células y, a continuación, se inyecta el núcleo de la célula somática que se desea clonar (donante), dentro del oocito previamente enucleado (receptor). El transplante de núcleos somáticos dentro de oocitos enucleados tiene como fin reproducir los procesos bioquímicos y fisiológicos que, de manera natural, se desencadenan durante la fertilización. Vale entonces, en este orden de ideas, establecer una comparación entre la fertilización y la clonación. Durante la fertilización, el primer evento que ocurre es la sincronización de los pronúcleos masculino y femenino. Cuando el espermatozoide atraviesa la membrana plasmática del oocito, su núcleo se encuentra en la etapa G0 del ciclo celular, mientras que el núcleo del oocito está detenido en la metafase de la segunda meiosis. Los oocitos recientemente ovulados presentan una elevada concentración de Factor Promotor de la Maduración (siglas en inglés MPF), una proteincinasa que induce la ruptura de la membrana nuclear y la condensación de los cromosomas, motivo por el cual pueden hacer su ingreso al núcleo ciertos factores citoplasmáticos que son necesarios para la replicación del ADN16. Los dos pronúcleos se llevan entonces a la etapa S del ciclo, caracterizada por una síntesis activa de ADN. La reanudación de la segunda meiosis ocurre como consecuencia del aumento cíclico de [Ca2+] intracelular, que provoca a su vez la inhibición de las dos subunidades constitutivas del MPF -ciclina B y cdc1- con el descenso consecuente en la actividad del MPF17,18. Cuando, en la clonación, se introduce el núcleo de la célula somática donante en la célula receptora u oocito, se efectúa un proceso similar de sincronización o reprogramación del ciclo celular, con la finalidad de que ocurra un acoplamiento fisiológico entre el núcleo y el citoplasma. La reprogramación del ciclo celular, o lo que es lo mismo, la reprogramación nuclear, son términos que describen los cambios en la actividad génica inducida experimentalmente por el transplante de un núcleo dentro de un medio citoplasmático diferente. La reprogramación es un requisito indispensable para el éxito del procedimiento, pues permite que la expresión génica de la célula somática sea la apropiada para un desarrollo embrionario normal13,19. Esta reprogramación se efectúa en el tiempo transcurrido entre la fusión núcleo-citoplasma y la activación del producto resultante19. Si se transplantan núcleos de células somáticas, parcial o totalmente diferenciadas, dentro de oocitos enucleados de anfibios o de mamíferos en metafase de la segunda meiosis, se podrán obtener blastocistos, a partir de los cuales se formará un amplio rango de tejidos y de tipos celulares20. El proceso de sincronización o reprogramación se efectúa mediante el estímulo de la entrada de Ca2+ al oocito, ya sea con un impulso eléctrico suave o con la utilización de ionomicina13,20 o puromicina19. El núcleo de la célula somática donante no debe haber iniciado o completado la replicación del ADN, para que el cigoto pueda tener una ploidía o complemento cromosómico normal. Si se activa el oocito, y se le permite entrar a su primer ciclo celular, la actividad del MPF caerá y no ocurrirá la ruptura de la membrana nuclear después de la transferencia del núcleo de la célula donante. Así las cosas, el núcleo determina si habrá o no replicación del ADN, de modo que pueda esperarse una ploidía normal -necesaria para un normal desarrollo- en todos los estados del ciclo celular de la célula donante16. Se considera entonces que el oocito es un receptor universal, pues no sólo provee un medio apropiado para el núcleo de la célula donante en cualquier estado del ciclo sino que, independientemente de éste, tiene la capacidad para actuar sobre la estructura y la función de la cromatina del núcleo somático, de modo que lo lleva de nuevo a un estado de totipotencialidad16,21. Al aplicar la técnica de la transferencia nuclear, no sólo se debe procurar mantener una ploidía normal en el producto formado, sino también evitar la destrucción de la cromatina. Por este motivo se han hecho investigaciones sobre la evaluación de la técnica, cuando se utilizan células donantes en diversas etapas del ciclo celular (G1, S, G2, M), y como células receptoras, oocitos en metafase de la segunda meiosis13,22-28. Como células somáticas donantes, al principio se utilizaron las células quiescentes en fase G0, que habían abandonado el ciclo celular en fase G1 y, que, por tanto, tenían un complemento cromosómico diploide. Como ejemplo de células en fase G0 se encuentran células del cúmulus, células de Sertoli y neuronas, aunque también se pueden obtener artificialmente en un medio de cultivo, mediante la deprivación de factores de crecimiento29. Aunque estas células son menos activas en la transcripción y, además contienen poblaciones distintas de ARN mensajero (ARNm) que las que se hallan en fase G1, se prefirieron, pues sus mismas características, de alguna manera facilitan la acción de los factores citoplasmáticos del oocito necesarios para modificar la expresión génica del embrión, y, por tanto, la reprogramación nuclear13,16. También se han utilizado células somáticas donantes en fases G125,28, G226 y M27 (Gráfica 1). Se ha obtenido una notable eficiencia en el desarrollo de los embriones así clonados hasta el estado de blastocisto, cuando el núcleo inyectado se encuentra en fase M22,23. En cualquier caso, una clonación exitosa requiere reprogramar el núcleo de la célula somática donante, debido a que la cromatina ha sufrido previamente transformaciones epigenéticas incompatibles con el desarrollo embrionario, que incluyen la desacetilación de las histonas y la metilación del ADN30-32. Recientemente, Hwang et al.19 obtuvieron células madre embrionarias a partir de blastocistos humanos. El éxito de su procedimiento radicó en que el tiempo destinado a la reprogramación celular fue de unas pocas horas, en contraste con la metodología utilizada en experimentos con otras especies de mamíferos33. Sin embargo, no siempre la reprogramación nuclear ocurre de modo satisfactorio. Así hay informes de hallazgos de embriones clonados a partir de células somáticas donantes, que fracasan en la reactivación de genes claves para el desarrollo embrionario normal31. En estos embriones se pueden expresar precozmente genes específicos de las células donantes34, y, adicionalmente, presentar patrones aberrantes de metilación del ADN en el trofoectodermo35,36, así como mosaicismo35. Por otra parte, las células somáticas, a lo largo de las sucesivas divisiones mitóticas sufridas in vivo, han perdido parte de los extremos de los cromosomas, denominados telómeros. Los telómeros son repeticiones cortas en conjunto (tándem) de ADN, que se replican mediante la acción de la enzima telomerasa. Se ha encontrado que variables como la longitud de los telómeros y la actividad de la telomerasa están significativamente disminuidas en las células somáticas donantes, cuando se comparan con las células madre embrionarias como fuente donante de núcleos. Debido a que los telómeros desempeñan un papel estabilizador del ADN, su acortamiento progresivo puede incidir, tanto en los procesos de envejecimiento celular, como en la aparición de anormalidades cromosómicas, espermatogénesis defectuosa, apoptosis aumentada y proliferación celular disminuida en médula ósea, bazo y testículo, en animales clonados a partir de células somáticas donantes21. Es claro entonces que el estado de diferenciación de la célula donante afecta directamente la eficiencia de la clonación, y en tal sentido se considera que las células madre embrionarias ofrecen ventajas cualitativas sobre las células somáticas como posibles donantes en la técnica de transferencia nuclear23,30,37. A pesar de los éxitos sucesivos obtenidos en la clonación de células adultas, la SCNT dista mucho aún de ser considerada una técnica eficaz. En primer lugar, muchos embriones clonados mueren inmediatamente después de la implantación, o bien, a lo largo del desarrollo prenatal38-40. También se ha observado, que si bien algunos individuos sobreviven hasta el término de la gestación, mueren prontamente debido a un amplio rango de diversas entidades35. Si logran sobrevivir, presentan una talla corporal y un tamaño de la placenta mayores que lo normal38,41-45, obesidad46, así como defectos en riñones, corazón, hígado, pulmones y cerebro47. Hasta hace poco tiempo se desconocía cuál era la razón biológica y cuáles los problemas técnicos causantes de tales fracasos48. En el caso de primates no humanos, como los monos Rhesus macacus, los intentos de clonación han fallado, debido a que, durante la enucleación de los oocitos no fertilizados -previa a la transferencia nuclear- se removían ciertas proteínas necesarias para la organización normal del huso mitótico, como las proteínas motoras de los microtúbulos y del centrosoma35,49. La dificultad fue en apariencia resuelta por Hwang y su equipo de colaboradores19, quienes recientemente obtuvieron blastocistos humanos. Ellos extrajeron el complejo ADN-huso mitótico inmediatamente después de la aparición del primer cuerpo polar, a través de un orificio en la zona pelúcida, en lugar de aspirarlo con una pipeta de vidrio, tal como es descrito por otros autores8. En especies de mamíferos como perros, conejos, cerdos, caballos, gatos y seres humanos, el éxito de la clonación ha sido altamente dependiente de la disponibilidad de tecnologías especie-específicas, que incluyen el cultivo, la activación, la micromanipulación y la transferencia de huevos y embriones a receptores19,50-55. Es así como en la aplicación de la SCNT en cerdos, si se efectúan simultáneamente los procesos de fusión núcleo-citoplasma y activación del producto, se obtienen resultados satisfactorios. En contraste, cuando se utiliza el mismo proceso en bovinos y en seres humanos, los porcentajes de fusión y de clivaje o segmentación son muy bajos y no se forman blastocistos. Por tanto en estas especies, parece ser necesario un mayor tiempo de reprogramación entre fusión y activación con el fin de obtener un número significativo de blastocistos. Por otro lado, el tipo de sustrato energético añadido al medio de cultivo (fructosa en lugar de glucosa) también parece ser un factor determinante para la formación de blastocistos en bovinos y en seres humanos19,56. 3. Paraclonación. Consiste en inyectar núcleos de células madre embrionarias en cultivo, dentro de oocitos enucleados y, a veces, de cigotos enucleados. Los blastómeros se obtienen a partir de varias fuentes como la masa celular interna o el trofoectodermo de embriones preimplantados y sus núcleos son transferidos dentro de oocitos enucleados6. Los individuos obtenidos son casi idénticos entre sí, aunque diferentes a los padres del embrión que aportó el núcleo transferido57. Se ha demostrado que la supervivencia de los clones formados a partir de células madre embrionarias en el instante del nacimiento o hasta la edad adulta, es 10 a 20 veces mayor que en los derivados de células somáticas23,30,37,43. Por otra parte, los clones derivados de células madre embrionarias tienen mejores posibilidades de reactivar completamente genes claves para el desarrollo embrionario -tales como oct4 y 10oct4- y así constituir una población de células verdaderamente totipotenciales19,23,31. Sin embargo, las células madre embrionarias, aunque requieren un menor grado de reprogramación que las células somáticas, presentan una elevada inestabilidad epigenética en cultivos in vitro. La inestabilidad se refleja en una expresión aberrante de la huella genética, de modo que, cuando estas células se utili-zan como donantes para la clonación reproductiva, el fenotipo fetal y placentario de los clones, exhibe graves patrones de anormalidad. Sin embargo, cuando se emplean como fuente de núcleos para la clonación terapéutica, en apariencia no ocurre este error, pues, durante la reprogramación, se seleccionan tan sólo las células competentes30,37,41. APLICACIONES DE LA CLONACIÓN MEDIANTE LA TÉCNICA DE TRANSFERENCIA NUCLEAR El estado del arte en la clonación ha permitido su empleo en dos direcciones claramente definidas: La clonación con fines reproductivos y la clonación con fines terapéuticos. La primera apunta a duplicar seres vivos completos, mientras que la segunda promete convertirse en una alternativa para prevenir y tratar ciertas enfermedades, así como para el reemplazo de tejidos y órganos lesionados (Gráfica 2). Clonación terapéutica. Las células madre o troncales son pues células pluripotenciales de gran tamaño que, después de experimentar un proceso de diferenciación, se especializan en una dirección funcional determinada, hacia una gran variedad de tipos celulares58. Las células madre se pueden obtener en dos formas diferentes: 1. A partir de células de la masa celular interna o del trofoectodermo de blastocistos clonados mediante la técnica de transferencia nuclear y cultivadas in vitro19. A las células somáticas donantes se las manipula para inducirlas a un proceso de diferenciación en tipos celulares determinados, que posteriormente se utilizarán en el tratamiento de ciertas enfermedades incurables. La ventaja de esta técnica radica en que las células madre embrionarias, una vez transplantadas, no provocan rechazo inmunológico, pues se comportan como injertos autólogos, gracias a que son genéticamente idénticas a las células del receptor o del paciente. El problema de los transplantes heterólogos, es el largo tiempo que toma el enfermo en aceptarlos, a pesar de que los órganos utilizados comparten su misma información genética, pues casi siempre provienen de miembros de la misma familia. Por otra parte, cuando se produce rechazo al tejido transplantado, la salud del individuo queda seriamente comprometida y hay la eventualidad que se debe pensar en un nuevo transplante. Algunos hallazgos recientes han revolucionado la biología de las células madre y han demostrado su potencial clínico para tratar diversas enfermedades, como trastornos neurodegenerativos, desórdenes sanguíneos y diabetes. Además, una estrategia válida para el manejo de desórdenes de origen genético conocido, como la anemia falciforme y la ß-talasemia, consiste en utilizar la técnica de transferencia nuclear, combinada con terapia génica y celular30,35. Los defensores de la clonación terapéutica de células humanas aducen que permite disponer de tejidos y de órganos viables, a partir de una fuente de ADN, sin que sea necesario traer un nuevo ser al mundo con el único fin de obtener un tejido59-61. Con la finalidad de asegurar la normalidad del clon y, por tanto de las células madre embrionarias, se propone una “prueba o test de seguridad” que seleccione y discrimine los embriones a los que se les permitirá progresar en su desarrollo. En teoría, este filtro se podría efectuar mediante la evaluación de los errores en la expresión y en la huella genética de los blastómeros en estado de preimplantación, pero no tiene en cuenta que se desconocen los efectos epigenéticos adversos que desencadenaría tal manipulación35. El asunto controversial es la conveniencia de crear embriones de seres humanos con la única finalidad de obtener líneas celulares, como también tejidos y nuevos órganos que se puedan utilizar en el reemplazo de los que se encuentran lesionados. Así se han levantado innumerables voces que de plano rechazan cualquier tipo de investigación que comprometa embriones de seres humanos. Argumentan que, para el cultivo de las células madre que forman parte del embrión, es necesario separarlas, y, por tanto destruir el paquete de células que se origina como producto de la fecundación. El meollo del asunto ético es “el sacrificio de embriones que sirven solamente como bancos de tejidos”59,60. Se ha cuestionado el uso de células madre embrionarias humanas con finalidades terapéuticas, no obstante sus encomiables objetivos, no sólo por reconocidos bioéticos sino también por organizaciones defensoras de los derechos humanos, con el argumento que se trata de un daño enorme a la vida del ser humano62,63. 2. A partir de ciertos órganos o tejidos de individuos postnatales, donde se ha demostrado que pueden persistir en forma indiferenciada. A la luz de conocimientos actuales de la ingeniería genética, estas células podrían ser transformadas en células madre pluripotenciales y cultivadas separadamente, de acuerdo a la necesidad del paciente48. Así, dentro de tejidos clásicamente considerados con un potencial regenerativo nulo, como el nervioso64,65 y el muscular66, se han identificado grupos de células madre, capaces de proliferar y de madurar hacia diferentes tipos celulares, tanto in vivo como in vitro. De la misma manera, en la zona subventricular del cerebro adulto, se han hallado células madre que eventualmente se podrían utilizar en terapias de reemplazo neuronal. Por otro lado, se observa una buena regeneración de tejidos como músculo esquelético lesionado, cuando se injertan mioblastos cultivados in vitro. Otra fuente prometedora de células pluripotenciales es la sangre del cordón umbilical. La muestra de sangre se obtiene en el momento del nacimiento, sin que el neonato ni su madre se vean afectados. La eficiencia del procedimiento es tal, que, a partir de un volumen de 20 ml de sangre del cordón, se obtienen hasta 4 millones de células madre67, que pueden crioconservarse por largo tiempo sin deterioro alguno, para ser utilizadas en transplantes68 y en procesos de terapia génica. La sangre del cordón umbilical también provee hematíes normales y leucocitos muy útiles en el tratamiento de la anemia falciforme y en la restauración del sistema inmunitario de los niños nacidos con inmunodeficiencia grave69,70. Si las células madre que provienen de determinados órganos de individuos adultos, se cultivan en condiciones apropiadas, son susceptibles de sufrir transdiferenciación. En 1999, un grupo de científicos italianos y canadienses demostraron la presencia, en personas adultas, de células madre nerviosas capaces de diferenciarse en células hematopoyéticas. Según sus hallazgos, la diferenciación de las células madre estaría condicionada por las señales que reciben del entorno donde se sitúan71. Igualmente se ha determinado que células madre nerviosas de ratones se pueden diferenciar hacia células hemáticas, como también las células hemáticas en células musculares esqueléticas72, o bien, en células de microglia o de astroglia73. Estos hechos sugirieron la posibilidad que células madre de la médula ósea fueran transplantadas con el fin de tratar enfermedades como distrofia muscular, mal de Parkinson, infarto de miocardio o falla hepática. Sin embargo no es claro si la aparente plasticidad de las células madre adultas examinadas se debe a las condiciones particulares en las que se cultivaron, a posible contaminación o a fusión celular. Además de esta incógnita que retrasa su empleo como donantes en la clonación, se ha observado que las células madre adultas presentan otra serie de desventajas con respecto de las células madre embrionarias. Tales desventajas incluyen no sólo dificultades en su aislamiento y cultivo, sino también una alta probabilidad de sufrir mutagénesis insercional y cáncer, como resultado de la introducción de transgenes retrovirales, necesarios para su manipulación genética74. En contraste, las células madre embrionarias se obtienen fácilmente a partir del embrión seleccionado, proliferan de modo indefinido en cultivo, y sus defectos genéticos se pueden reparar mediante procesos de recombinación homóloga75. Recientemente se obtuvieron células madre adultas derivadas de tejido mesenquimatoso de médula ósea en ratas, ratones, otros animales y seres humanos76, que sufrieron una rediferenciación exitosa hacia células de las tres capas germinativas (ecto, meso y endodermo), cuando se transfirieron sus núcleos dentro de blastocistos. Hay otras fuentes, también cuestionadas, que ofrecen disponibilidad de células madre pluripotenciales humanas, que se pueden diferenciar y cultivar separadamente: Son los embriones desechados durante el procedimiento de fertilización in vitro (IVF)4,77. Clonación reproductiva. Inicialmente, el proceso es igual al que se efectúa durante la clonación terapéutica. La diferencia aparece con posterioridad a la fusión del núcleo de la célula donante con el oocito enucleado, pues el cigoto se debe implantar en un útero, donde se desarrollará hasta formar un individuo réplica del donante. Dicen sus defensores, que esta técnica podría ser una alternativa para las parejas infértiles o, para las que tienen hijos con enfermedades genéticas o, que potencialmente podrían engendrarlos. Sin embargo, los detractores consideran que es un procedimiento “biológicamente incierto”78 debido al poco éxito alcanzado en la clonación de primates no humanos35, amén de poco práctico, pues tendrían que fertilizarse muchas madres sustitutas con el fin de obtener un solo nacimiento exitoso79. Además, la necesidad médica para efectuar una clonación reproductiva es mínima, si se tiene en cuenta el porcentaje relativamente bajo (1%) de parejas infértiles78. En favor de la clonación con fines reproductivos, se ha esgrimido que representa una opción para las “parejas de homosexuales” que desean tener hijos propios. Tal es el caso de las parejas femeninas que no desean tener un bebé a partir del esperma de alguien desconocido, como ocurre con los procedimientos de reproducción asistida (IVF). También podría ser una luz de esperanza para los padres que han perdido a un hijo. Sin embargo, ninguno de estos argumentos ha sido acogido por las organizaciones internacionales de bioética o por los científicos geneticistas. Así, se ha afirmado que la clonación no puede hacer de nadie un ser inmortal, porque el clon es una persona diferente80, y, aún se debate si el clon tendría los mismos derechos que cualquier otro ser humano. La verdad es que si la clonación con fines reproductivos estuviera permitida, de seguro la practicarían preferentemente las parejas infértiles, de manera semejante a lo que ocurre con los procedimientos de IVF, y no por individuos que motu proprio decidan reproducirse mediante este procedimiento. Por otro lado, si alguien desea clonar a una persona ya fallecida, debería haber tenido la precaución de obtener células suyas en vida, si se considera que la supervivencia celular postmortem es sólo de unas dos horas. Además, debería tener un consentimiento certificado del individuo donante. Otras razones éticas, como la identidad del clon, suscitan cuestionamientos, hipotéticamente aun por parte del mismo clon. En 1970 Hans Jonas, filósofo inglés -según Annas60- explicó que “la clonación es un crimen contra el mismo ‘clon’ pues lo priva de sus derechos de conocer la verdad acerca de su existencia.” En otras palabras, el clon estaría sujeto a la versión que el donante quiera suministrarle sobre el origen de su existencia y su legitimación como ser humano. En contraste, en el campo de la clonación animal con fines reproductivos, no se han generado tales controversias, pues existe consenso sobre las bondades de efectuar el procedimiento en animales de granja, con la finalidad de preservar el genoma de los mejores ejemplares. Las características genéticas más valoradas son, entre otras, un crecimiento rápido, resistencia a enfermedades, una elevada producción de leche o de lana de alta calidad. Si se toma la decisión de clonar animales, no sólo es factible predecir con facilidad sus características, sino también aumentar el bienestar de la población, mediante el aumento de la resistencia a ciertas entidades. Aunque la clonación utilizada en este contexto reduciría la diversidad, por otro lado ayudaría a esparcir mejores genes. La clonación reproductiva también permite preservar especies exóticas o que se encuentren en peligro de extinción7. CONCLUSIONES Y PERSPECTIVAS La aplicación de la clonación mediante la técnica de transferencia nuclear, no obstante las pocas modificaciones experimentadas en los últimos 10 años, ha permitido un mejor entendimiento de los mecanismos que regulan el desarrollo. Una consecuencia trascendental de lo anterior es la revelación de nuevos prospectos que se refieren a temas de estudio nunca antes imaginados, así como de aplicarlos tanto a la medicina como a las ciencias agropecuarias. Aunque la eficiencia de las técnicas empleadas hasta la fecha ha sido baja, su potencialidad es enorme, de modo que se espera que dentro de muy poco tiempo se pueda utilizar comercialmente, con el fin de incrementar, tanto la productividad como el bienestar de las poblaciones de animales de granja. Asimismo serán una realidad los animales clonados y manipulados genéticamente mediante la introducción de genes humanos, que se utilizarán para producir proteínas humanas, cuya deficiencia causa serias enfermedades como la fibrosis quística y la hemofilia. El futuro de la clonación con fines terapéuticos es prometedor, pues se podrán efectuar trasplantes de tejidos sanos en personas aquejadas por diversos tipos de enfermedades. Así los científicos esperan producir órganos completos en el laboratorio, para reemplazar los que se encuentran lesionados en pacientes con enfermedades devastadoras como cáncer, dolencias cardíacas, diabetes y mal de Parkinson. A este respecto, el debate ético no está centrado en sus loables propósitos, sino en la conveniencia de utilizar células madre de embriones de seres humanos versus células madre obtenidas a partir de individuos adultos. Sobre la clonación con fines reproductivos se ciernen serias incógnitas, debido a que casi todas las causas de las complicaciones vistas, permanecen en el misterio. En la práctica, su aplicación en el ser humano ha sido vetada no solo por eminentes científicos, sino por la gran mayoría de las organizaciones defensoras de los derechos humanos, que han expresado con claridad su rechazo al procedimiento, debido a convicciones que van desde lo social y ético, hasta lo religioso. En su Declaración Universal sobre el Genoma Humano y los Derechos Humanos81 del año 1997, en el artículo 11, la Conferencia General de la Organización de las Naciones Unidas para la Educación, la Ciencia y la Cultura (UNESCO), declara: “No deben permitirse las prácticas que sean contrarias a la dignidad humana, como la clonación con fines de reproducción de seres humanos. Se invita a los Estados y a las organizaciones internacionales competentes a que cooperen para identificar estas prácticas y a que adopten en el plano nacional o internacional las medidas que corresponda, para asegurar que se respetan los principios enunciados en la presente Declaración”. Por su parte, Wilmut et al.8, científicos del Roslin Institute que lideraron el equipo responsable de la clonación de la oveja Dolly, afirman: “Aunque parece técnicamente posible la clonación humana, no se debería intentar siguiera, pues es una aberración carente de utilidad clínica”. En concordancia con las declaraciones anteriores, un número significativo de países miembros de la Organización de las Naciones Unidas (ONU) ha venido propiciando el debate tendiente a la adopción de una convención internacional que regule las investigaciones sobre clonación humana. Es así como dentro del marco del VI comité de la 58° Asamblea General, el día 6 de noviembre de 2003, se presentaron dos propuestas: la primera, que abogaba por una prohibición total de las investigaciones sobre clonación humana, tanto reproductiva como terapéutica, avalada por un numeroso grupo de países liderado por los Estados Unidos de América, Costa Rica y España. La segunda, aunque también vetaba la clonación humana con fines reproductivos, hacía énfasis sobre la necesidad de investigar con células humanas clonadas, con miras a su posible utilización terapéutica. Esta última propuesta fue presentada por un grupo de países entre los que estaban Gran Bretaña, Bélgica y Singapur. No obstante la aparente claridad de las diferentes posturas, en el transcurso del debate se hizo evidente una gran polarización, que llevó a que los países pertenecientes a la Organización de la Conferencia Islámica (OIC), representados por Irán, promovieran una propuesta alterna que postergaba el debate hasta el año 2005. El objetivo de esta moción, que finalmente fue aprobada, era el de permitir que en el lapso previsto hubiese una mayor posibilidad de demostrar hasta qué punto son necesarias las células humanas clonadas en el tratamiento de enfermedades degenerativas82. Consciente de la importancia de adoptar con prontitud una posición definida al respecto, la Asamblea General de las Naciones Unidas, en encuentro efectuado el día 9 de diciembre de 2003, anuló la votación de su comité legal que postergaba por 2 años el debate, y, en consecuencia, decidió incluir el tema de la clonación humana con fines reproductivos en su agenda para la sesión 59 del año 2004. Igualmente instó a los países miembros a que una vez sea aprobada la convención, desarrollen una legislación coherente que prohiba cualquier tipo de investigación, experimentación, desarrollo o aplicación de técnicas orientadas a la clonación humana con fines reproductivos dentro de sus territorios. Por tanto, una legislación nacional tendrá que seguir a la resolución de la ONU, si se tiene en cuenta la no obligatoriedad de acatar sus resoluciones83. Aunque muy seguramente la prohibición de clonar seres humanos será adoptada internacionalmente por convención de la ONU durante el transcurso del año 2004, la radicalización de las diferentes posiciones en torno a la clonación humana con fines terapéuticos, no permitirá que exista consenso a su alrededor, al menos en el corto plazo. Así por ejemplo, se habla de la posibilidad de que los embriones utilizados como fuente de células madre provengan en su mayoría de los países en vía de desarrollo82 donde, en este orden de ideas, no sería inusual que un comercio ilegal de embriones y de tejidos embrionarios hiciera su aparición. REFERENCIAS
Copyright 2004 - Revista Colombia Médica The following images related to this document are available:Photo images[rc04017g2.jpg] [rc04017g1.jpg] |
| |||||||||
{kind=link}
{kind=link}