 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Revista Colombia Médica, Vol. 35, No. 3, 2004, pp. 150-160 La genómica nutricional: un nuevo paradigma de la investigación de la nutrición humana Felipe García-Vallejo, Ph.D. Profesor Titular, Director del Laboratorio de Biología Molecular y Patogénesis, Departamento de Ciencias Fisiológicas, Facultad de Salud, Universidad del Valle, Cali Recibido para publicación
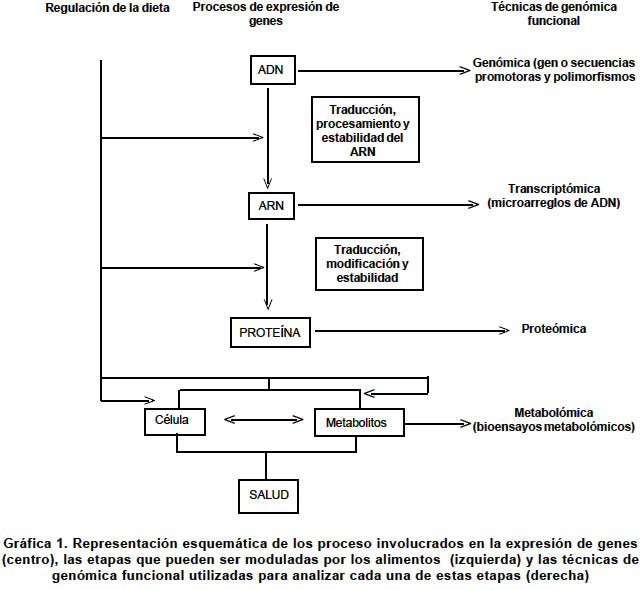
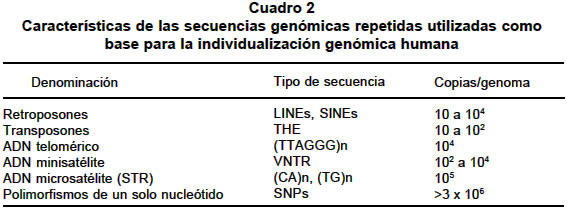
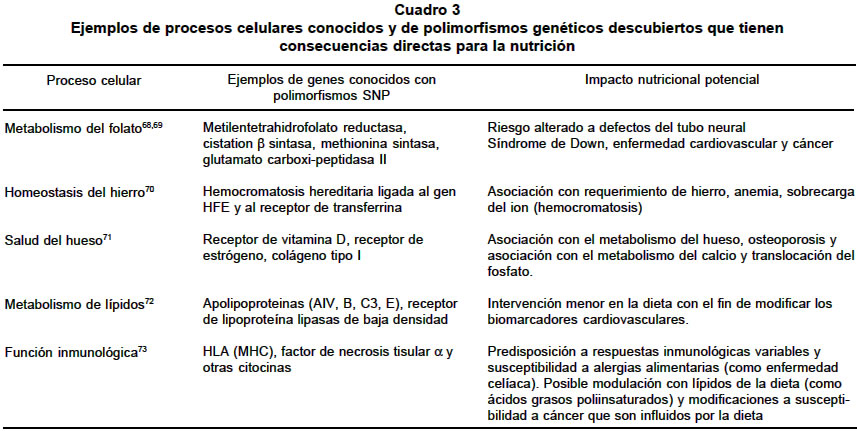
septiembre 3, 2003 Code Number: rc04025 RESUMEN La relación entre la dieta y la salud es un hecho que ha sido ampliamente demostrado; sin embargo, existe un interés creciente en cuáles de los componentes de la dieta son biológicamente activos y cómo ellos ejercen su efecto funcional. Estas preguntas han sido ejes centrales del desarrollo de la genómica nutricional. Así, la genómica nutricional es la aplicación de las tecnologías utilizadas en la funcional para entender, a nivel genómico, el efecto que tienen los alimentos. Una de las preguntas más comunes en nutrición es ¿Qué y cómo comer? La aceptada individualidad metabólica que se ha predicado en los últimos años, impide que se pueda dar una respuesta general. Actualmente se sabe que el genoma es individualizado y que la expresión de su contenido informacional es un universo único, que depende no sólo de factores exógenos sino que es modulado de manera muy compleja por múltiples factores endógenos. Es pues un reto de conocimiento el comenzar a definir cuáles son las variables que intervienen para poder tener una dieta acorde con cada individuo. En este sentido, la genómica nutricional (o nutrigenómica), plantea el estudio del efecto de los alimentos a nivel molecular y genético. En tal contexto, el proyecto del genoma humano provee herramientas para poder entenderla y se convierte en un nuevo paradigma de la investigación en nutrición humana. Palabras clave: Nutrigenómica. Genómica. Genoma humano. Genómica funcional. Transcriptómica. Salud pública. SUMMARY The link between diet and health is well established, however, renewed interest in which dietary components are biologically active and how they exert their functional effects is being fuelled by the development of nutritional genomics. Nutritional genomics is the application of high throughput functional genomic technologies in nutrition research. One of the most common questions in nutrition is: What to eat and how do it? The accepted metabolic individuality that has been proposed during the last years permitted to address these and other questions. Actually we know that the genome is individualized and the expression of its informational background is also unique; however, it is influenced by several exogenous factors and internally is controlled by several regulating factors that form complex nets of genes which interact among them. In this sense is a real challenge to explore what are the variables that play fundamental roles in the expression of the informational content of genome. Nutritional genomics introduce to the analysis of the effects of foods on the expression at molecular and gene levels. Thereby the knowledge of the human genome project is giving tools to enable us to understand better many biological aspects of the nutrition promoting the development of a new paradigm in nutritional research. Key words: Nutrigenomics. Genomics. Human genome. Funtional genomics. Transcriptomics. Public health. Las ciencias de la nutrición están descubriendo las que han sido denominadas ciencias “omicas”. Impulsadas por las recientes revelaciones del proyecto genoma humano y los desarrollos tecnológicos asociados, el genotipado, la transcriptómica, la proteómica y la metabolómica están disponibles ahora para ser utilizadas en la investigación en nutrición (Cuadro 1). En el futuro se verá la utilización de nuevas herramientas para la selección de nutrientes bioactivos, nuevos marcadores para definir in vivo la eficacia de los nutrientes además de un mejor conocimiento de la influencia de los polimorfismos genéticos sobre el metabolismo de los nutrientes. La genómica tuvo como base de su desarrollo la secuenciación de los más de 3 mil millones de nucleótidos del ADN humano; este logro ha permitido iniciar la identificación del conjunto de genes humanos y la asignación de la función biológica de estos1,2. A pesar de la complejidad inherente del genoma, la invención de unas pocas herramientas moleculares permitió el avance arrollador de la genómica en muchos campos de la investigación humana. Es razonable asumir que en un futuro no muy lejano, se puedan identificar las causas de la mayoría de las enfermedades genéticas3,4. El poder que emerge del genoma es que la identificación de las bases genéticas de las enfermedades hereditarias dará una estrategia lógica para tratar de manera individualizada las personas afectadas5. Más allá de la aplicación sobre el tratamiento de las enfermedades de base genética, existe un interés cada vez más creciente por definir cómo los genes interactúan con elementos de la dieta humana modificando el metabolismo celular y generando cambios en los perfiles metabólicos que pueden estar asociados con la susceptibilidad y riesgo a desarrollar enfermedades comúnes en las sociedades tecnológicamente avanzadas6-10. Los metabolitos son moléculas cuantificables que reflejan el fenotipo celular; sin embargo, la biotecnología moderna y la bioinformática han acometido una tarea de identificación de estas moléculas que es tan compleja como el mismo proyecto genoma humano11,12. Es necesario el entendimiento de los múltiples factores que influyen la salud humana y cuyo conocimiento se incrementará con la cuantificación de la acción de los metabolitos en cada individuo13. En este contexto, las bases de datos sobre el tipo y la concentración de metabolitos celulares serán un recurso predictivo para cuantificar las relaciones entre estos y la salud. Además, la informática de los metabolitos o metabolómica será el paso lógico siguiente para entender el papel de la nutrición en la modificación del metabolismo y definitivamente en la promoción de la salud14,15. No hay duda de que el mejoramiento de las propiedades nutricionales de los alimentos tanto de origen vegetal como animal y microbiano, será de considerable valor para la salud pública y para la industria de la nutrición16,17. Sin embargo, no hay todavía consensos sobre cómo alcanzar estas metas, particularmente sobre cómo identificar cuál molécula blanco específica se debe cambiar. Esta incertidumbre se genera irónicamente del limitado conocimiento tanto de la genética de las fuentes biológica de los alimentos como de los humanos, pero además de una vieja y muy considerada pregunta ¿Qué es nutritivo? Uno de los objetivos principales de la agricultura de plantas como fuente de alimentación, es producir cambios en los cultivos y los alimentos derivados que incrementen benéficamente la salud de todos los humanos18. En este sentido el desarrollo biotecnológico de plantas comestibles ha alcanzado niveles de calidad que permiten su utilización para una dieta nutricionalmente balanceada. Sin embargo, el enriquecimiento de fuentes alimentarias con dosis elevadas de ciertos metabolitos, bajo la creencia de que las altas concentraciones de ellos pueden jugar un papel benéfico en la persona, también ha tenido efectos negativos en la salud19. Se ha demostrado que niveles elevados de b-carotenos en fuentes vegetales, los cuales inicialmente se pensó podían ser protectores de procesos cancerosos, en algunos casos aumentaron el riesgo a desarrollar cáncer de pulmón en individuos fumadores20,21. LA VISIÓN MODERNA DEL GENOMA El genoma es el contenido total del material genético de un organismo, empacado en un juego completo de cromosomas. El genoma es el material portador de la herencia, es decir, es el que determina la transferencia de los caracteres hereditarios del progenitor a su progenie20. La disponibilidad de las secuencias completas del genoma humano y de otros organismos supone un cambio estratégico en el abordaje de los problemas biológicos1,2. Con la casi totalidad de los genes en la mano, la aproximación reduccionista convencional de escudriñar gen por gen puede complementarse por una aproximación más global o integradora para considerar todos los genes a la vez, sobre la base de que son las intrincadas redes de interacciones entre proteínas y moléculas de ácido ribonucleico (ARN) las que regulan los procesos biológicos, más que proteínas aisladas actuando en solitario. Tales aproximaciones, ayudan a comprender los mecanismos moleculares involucrados en las enfermedades humanas. Los proyectos de secuenciación de genomas se asocian cada vez con mayor frecuencia, con el reto de comprender la función de los genes identificados en proyectos de secuenciación a gran escala21. La “era postgenómica” ha sido declarada oficialmente abierta y ello incluye la descripción tanto de cada una de las proteínas expresadas por sus genes respectivos (proteómica)22, como del conjunto interactivo de las proteínas involucradas en un proceso o módulo biológico en particular (proteómica funcional)23. La complejidad inherente del tratamiento simultáneo de cientos de proteínas para formular problemas biológicos integrados exige disponer de mapas informativos cuya complejidad funcional es cada día mayor24 (Gráfica 1). En la actualidad existe un gran número de tecnologías que conforman la base experimental de la genómica nutricional (Gráfica 1). Sin embargo, muchas de ellas todavía no se han impulsado como pruebas en la ciencia de la nutrición; esto no significa que su gran potencial permita su adopción rápida en disciplinas donde la genómica nutricional tiene un efecto medible como son la farmacéutica, la toxicología, la investigación clínica y la farmacogenómica. El mayor reto de la genómica nutricional está en el diseño de estudios significantes que apliquen estas metodologías, el desarrollo de investigaciones capaces de descifrar las complejas interacciones entre las diferencias genéticas individuales, la predisposición a las enfermedades, el componente de interacción genómica además de tratar de integrar los grupos de estudios que se han realizado en el pasado. La bioinformática, una nueva aplicación del genoma. Los organismos pueden concebirse como sistemas de módulos moleculares de expresión de genes, cada uno de ellos, responsable de una determinada función biológica. La función de los módulos puede ser básica como la síntesis de una proteína, la regulación del ciclo celular o la maquinaria de reparación del ácido desoxirribonucleico (ADN), o compleja como las memorias a largo plazo e inmunológica. Uno de los principales retos de la era postgenómica es que la mayoría de los genes identificados en las secuencias genómicas completas deben ser adscritos a un determinado módulo funcional. Esto genera la pregunta de si cada módulo estudiado incluye más genes de los supuestamente involucrados o si existen más módulos que deban ser descubiertos25,26. En cualquier caso, la estrategia de partida asume que los genes copartícipes de un determinado módulo funcional deben compartir propiedades comunes (interactoma). Así, los mapas de transcriptomas consistirán en un conjunto de “grupos de expresión” o de genes corregulados, mientras que el interactoma contempla redes de proteínas interactivas (grupos de interacción). Una de las principales limitaciones de los mapas funcionales es que sólo ofrecen una aproximación muy general de los genes pertenecientes a un módulo; hoy sólo somos capaces de generar conclusiones sobre la función de unas pocas proteínas. No cabe duda de que las restricciones inherentes a los mapas funcionales se irán disipando conforme más tipos de mapas vayan integrando conjuntos solapantes de características funcionales. De este modo, uno de los retos más evidentes de la era postgenómica es aprender cómo puede integrarse la información funcional aportada por los distintos tipos de mapas; es una empresa que sólo se resolverá mediante la colaboración computacional. La bioinformática se asienta, cada vez con mayor solidez, como la herramienta clave de la empresa biológica del futuro. Otro reto es aprender cómo la información funcional de los diferentes mapas puede integrarse en un atlas biológico. Generaciones de bioinformáticos se ocuparán de desarrollar la infraestructura de los mapas funcionales y la arquitectura de los atlas biológicos. Efectos de la alimentación moderna. Los alimentos que comemos contienen miles de sustancias biológicamente activas, muchas de las cuales pueden tener un potencial para ejercer un efecto benéfico para la salud3,19. Muchos compuestos derivados de fuentes alimentarias tales como el sulforofano, la curcumina, el licopenos y los polifenoles del té, se convierten en los más promisorios agentes quimiopreventivos conocidos27. El impacto de los componentes biológicos activos de la dieta es muy poco conocido y el entendimiento de sus mecanismos de acción es aún más limitado28,29. Muchos de los datos que actualmente se dispone se han derivado de estudios in vitro con moléculas altamente puras en formas y concentraciones a las cuales los tejidos del cuerpo nunca serían expuestos. Si bien este trabajo es un punto de partida, se requiere obtener un conocimiento integral en sistemas modelo fisiológicamente más reales en los que se incluyen la caracterización del efecto de la tasa de absorción, la dispersión en el tejido y los sitios blanco de compuestos metabólicamente importantes además de la de desarrollar estudios en tiempo real del efecto dosis respuesta30,31. Clásicamente la nutrición se ha centrado en la investigación reducida a unos pocos tejidos para estimar el riesgo de individuos a desarrollar ciertas enfermedades por la acción de los alimentos; día a día cobra más importancia la evaluación integral de estos alimentos sobre el genoma teniendo en cuenta que existe una heterogeneidad genética dentro de la especie humana32,33. El patrón de enfermedad como humanidad se ha modificado enormemente desde hace unos cien años debido a los cambios en los hábitos alimenticios y sobre todo, a la manipulación de los alimentos34-37. Los alimentos procesados, refinados, que constituyen un alto porcentaje en la dieta, carecen de muchos nutrientes que deberían tener además, generan sustancias que pueden ser potencialmente nocivas para la salud38-40. El cocimiento, enlatado, almacenamiento, congelamiento y demás métodos de manejo de los alimentos, en la mayoría de los casos les quitan muchos de sus nutrientes. P.e., todas las enzimas (existen más de 3,700 de ellas) son sensibles al calor. Se destruyen entre los 60º y 80º C. Lo mismo sucede con muchas vitaminas y algunos aminoácidos41,24. Eso significa que los alimentos ya no tendrán el valor nutritivo original que normalmente tienen en estado no procesado. Así, al cocer un alimento, se destruyen muchos nutrientes. ¿Ahora qué pasa cuando se frien?43,44 La propia naturaleza da la opción del germinado en el caso de las semillas difíciles de digerir en estado crudo. En la actualidad, la medicina ortodoxa no le ofrece mucho a aquellos que sufren de ciertas enfermedades genéticas. Por otro lado, la terapia bioquímica nutricional es tal vez más poderosa que nunca cuando se utiliza para prevenir el desarrollo de las enfermedades. Hace algunos años, Williams45 propuso el concepto de la individualidad bioquímica para referirse a que cada individuo tiene un metabolismo propio y diferente al de los demás. Todo este conocimiento permite ahora a los médicos, ecólogos y nutricionistas encontrarse en una situación en la que pueden ayudar a personas con predisposiciones genéticas a prevenir la enfermedad en una forma efectiva. La dieta puede cambiar el estado de ánimo. Puede ayudar a mejorar la memoria y retardar el envejecimiento. También el cambio de estilo de alimentación puede curarnos de algunas enfermedades; pero el punto más importante es, tal vez, que la alimentación puede prevenir muchas enfermedades incluyendo el cáncer46,47. El diseño de las dietas: genoma vs. ambiente. Las frutas que se consumen en la dieta diaria pueden ser sustituidas por extraños compuestos y aminoácidos que potencialmente evitan enfermedades. Ese es uno de los objetos de la genómica nutricional48. Esta nueva frontera de la investigación molecular estudia el efecto de la nutrición en el nivel genético. Es posible ahora olvidarse de la “dosis diaria recomendada” y todas las normas ideadas para la población en general. Las nuevas investigaciones aportarán dietas “hechas a la medida” para cada persona, que estén acordes con su constitución genética49,50. Desde hace mucho tiempo se viene estudiando la relación que existe entre la dieta y el metabolismo. ¿Cómo es que algunas personas que consumen muchas grasas no padecen enfermedades cardiovasculares? ¿Por qué razón aquellos que no comen muchas grasas tienen un nivel muy alto de colesterol? Estas son algunas de las preguntas que pretende contestar la genómica nutricional. La individualidad del genoma y sus consecuencias en el metabolismo. Varias décadas de investigación en genética humana proporcionaron el conocimiento suficiente para poder particularizar el genoma de cada individuo. El conocimiento del alto grado de repetición del genoma humano ha permitido caracterizar muchas de las secuencias repetidas, algunas de las cuales se han empleado como instrumentos para individualizar el genoma humano (Cuadro 2). Jeffreys51,52 demostró que la distribución y el número de una serie de secuencias repetidas denominadas minisatélites era una marca personalizada. Se acuñó el término “huella dactilar genética” (“DNA fingerprinting”) para describir la individualidad genómica de los humanos como una consecuencia de la historia evolutiva de las poblaciones. Los minisatélites o VNTR (“Variable Nucleotide Tandem Repeats”) son loci que corresponden a secuencias de ADN de unas pocas decenas de nucleótidos repetidas en tandem. El número de dichas repeticiones varía de cromosoma a cromosoma, de tal manera que en un cromosoma el número de repeticiones en tandem puede ser de 10, en otro de 15, en otro de 22, etc. La singularidad más especial de este tipo de polimorfismos está en que cada loci puede presentar muchos alelos distintos (tantos como repeticiones), sin embargo, presentan el inconveniente de no estar distribuidos por todo el genoma y por tanto sólo pueden ser utilizados en el diagnóstico de un número muy reducido de enfermedades. Los VNTR-minisatélites han encontrado su máxima aplicación en la determinación de la paternidad y en los protocolos de identificación genética en el ámbito judicial53,54. El desarrollo del proyecto genoma humano reveló otros tipos de individualidad genética cuya resolución era mayor que los minisatélites. Así, las repeticiones trinucleotídicas, STR o microsatélites y los polimorfismos de un solo nucleótido55 o SNPs recientemente analizados56, han mostrado una vez más la individualidad del genoma de los humanos. Si bien es cierto estas marcas genéticas son en principio neutrales ya se comienza a entender, sobre todo en los SNPs, que pueden conferir alguna ventaja adaptativa. Los microsatélites (STRs) son secuencias de ADN repetitivas simples constituidas por unidades de repetición de 2 a 7 pares de bases (pb) y de aproximadamente 100-400 pb de longitud total, distribuídas por el genoma de las células eucariotas. Su abundancia y su naturaleza polimórfica los convierte en marcadores útiles para diversos tipos de análisis. Cada microsatélite específico puede ser amplificado por PCR (reacción en cadena de la polimerasa) y visualizado por distintas técnicas57-59. En promedio hay cerca de 3 millones de posiciones dentro de las cuales el genoma humano puede variar en un nucleótido. A estas variaciones se les conoce como SNPs o polimorfismos de un solo nucleótido y determinan la individualidad del genoma humano60-67. No todos los SNPs o polimorfismos individuales, son causantes de enfermedades; sin embargo, algunos SNPs confieren riesgo genético para enfermedades comunes68-73, porque la expresión de la variante de estos genes codifica para proteínas cuya función no corresponde a la norma. Algunos de estos polimorfismos se localizan en genes que codifican por enzimas cuya variación polimórfica puede asociarse con enzimas cuya actividad puede ser modulada por componentes de la dieta (Cuadro 3). Sin embargo, aunque la individualización genética es una herramienta muy poderosa en el estudio de los genomas, no proporciona información con respecto a su función tanto en el tejido como en el contexto de la interacción con el ambiente; esta limitación comienza a ser objeto de estudio y se espera que en un futuro no muy lejano pueda dar información fundamental para entender el efecto de los alimentos en la expresión de los genomas (genómica funcional). La genómica funcional. La genómica nutricional ha demostrado que los alimentos juegan un papel fundamental modulando el balance homeostático de múltiples procesos fisiológicos que están directamente asociados con la expresión de los genes. Ahora, como resultado del proyecto genoma humano, hay los instrumentos necesarios para estudiar en detalle cómo los nutrientes modulan la expresión del genoma del individuo. Es importante en este punto, definir dos de los componentes del genoma humano:
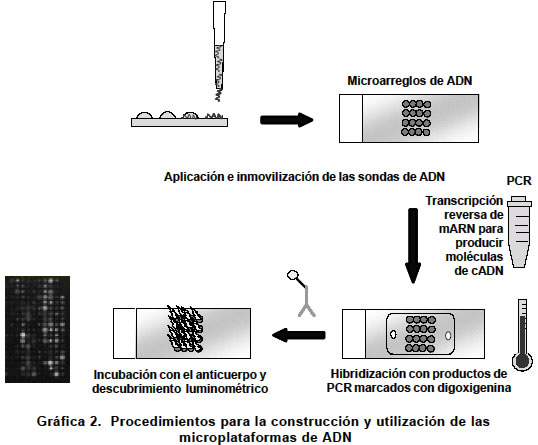
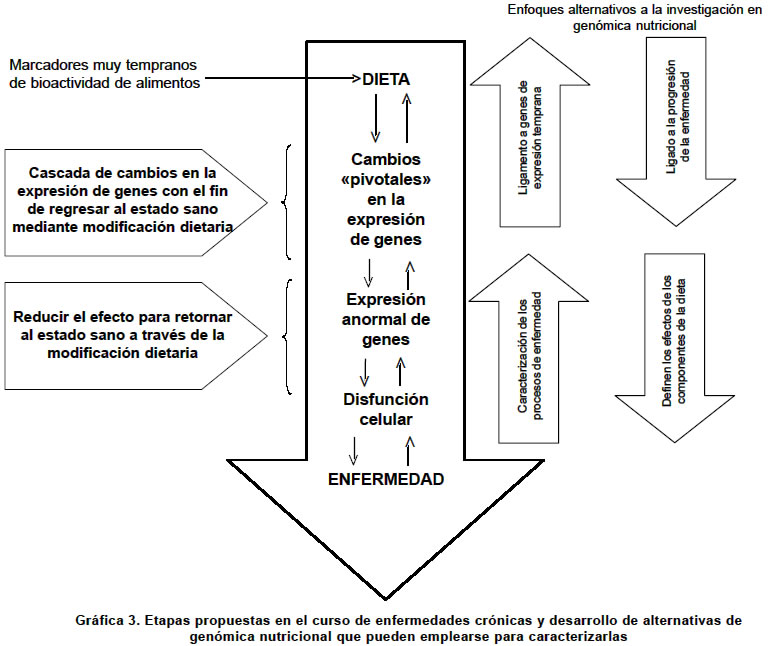
Las microplataformas de ADN o chip de ADN. Un chip de ADN es una microplataforma que contiene unidas múltiples secuencias de ADN que potencialmente sirven de sondas para descubrir, mediante hibridización de ácidos nucleicos, los distintos cADN que se han sintetizado in vitro a partir de poblaciones de mARN extraídas de una célula (Gráfica 2). Como su nombre lo infiere, este arreglo no ocupa más de unos cuantos centímetros y permite colocar de manera individual aproximadamente 30,000 tipos diferentes de sondas incluyendo cADNs y ESTs (secuencias expresadas con marcas), que pueden identificar un número equivalente de ARNs mensajeros o secuencias de nucleótidos expresadas en una célula. Lo que se obtiene con esta tecnología es un perfil individual de la expresión de los genes, la cual es una radiografía del funcionamiento del genoma. En la actualidad existen dos formatos de microarreglos de ADN que se diferencian entre si en el tipo de ácido nucleico empleado como blanco de hibridización: Formato I (microplataforma). Contiene sondas de cADN (500 a 5,000 bases en extensión) que se inmovilizan sobre una superficie sólida (vidrio) utilizando un robot aplicador, separado o en una mezcla, se pueden incluir entre 300 y 6,000 blancos de hibridización. Formato II (chip de ADN). Contiene oligonucleótidos (20 a 80 monómeros) o péptidos o PNA (sondas de ácidos nucleicos) que se sintetizan in situ (sobre el chip) o por síntesis convencional usando un inmovilizador. Para este propósito se utilizan técnicas litográficas de alta resolución. En este formato se pueden incluir entre 20 y 30,000 blancos de hibridización con lo que se expande de forma espectacular el rango de determinación de la expresión de genes en células específicas. Microplataformas de ADN de alta densidad. La clave que caracteriza a todos los microarreglos de ADN es que moléculas de ácidos nucleicos que están marcadas, hibridizan en solución con alta especificidad a secuencias inmovilizadas en un sustrato sólido. Así se facilita la medición cuantitativa en paralelo de muchas secuencias de ADN presentes en una mezcla compleja. Aunque se han desarrollado muchos métodos de microplataformas de ADN, actualmente existen dos que han sido perfeccionados y son los de uso común en la actualidad. En uno de ellos, los microarreglos de ADN se construyen mediante unión física de fragmentos de ADN procedecentes de clones originarios de bibliotecas génicas o a partir de productos de la reacción en cadena de la polimerasa. Utilizando un robot capaz de arreglar e imprimir capilarmente secuencias de ADN se pueden fijar hasta 23,000 elementos en una placa de microscopio. Por el otro método, los arreglos se construyen sintetizando oligonucleótidos de cadena única in situ empleando técnicas fotolitográficas. El revelado de las secuencias hibridizadas se efectúa por lo general mediante la emisión fluorescente de los híbridos y permite su cuantificación en un citodensitómetro. Las ventajas del primer método son su bajo costo y su flexibilidad; además, la secuencia primaria no se requiere para imprimir el elemento de ADN. En el segundo método se pueden barrer hasta 28,000 características en una placa de 1.28 x 1.28 cm aumentado su poder de resolución. Aunque todavía existen algunos inconvenientes tecnológicos, los microarreglos de ADN ya son una herramienta poderosa aplicada en la genómica nutricional74. La genómica nutricional funcional es una realidad. Aunque la individualización genómica permite enfocar la potencialidad del genoma de un individuo frente a una determinada dieta, la pregunta que surge es ¿Cómo estudiar la compleja trama de interacciones a nivel génico que permiten la expresión del genoma? En la última década del siglo XX, el desarrollo de la nanotecnología permitió la creación y perfeccionamiento del famoso chip de ADN como un instrumento que permitió disectar de manera muy fina la expresión, en forma de mARNs de los genes de una célula, dando como resultado un perfil genómico (Gráfica 2). Trabajos recientes75 han mostrado que la variación en las condiciones nutricionales produce una expresión global diferencial de los genes de una célula (cambio en el perfil genómico). Se ha demostrado que en el hígado de individuos sometidos a restricción calórica (RC) se observa una expresión de genes que es opuesta a aquellos que se expresan en el envejecimiento. El grupo de genómica nutricional del Departamento de Bioquímica de la Universidad de California liderado por el Dr. Stephen Spindler ha demostrado que durante el envejecimiento de la célula hepática de ratones se expresan genes asociados con la inflamación, el estrés celular, una disminuida capacidad para la apoptosis, el metabolismo xenobiótico, el ciclo celular normal y la replicación del ADN76. En contraste, la restricción calórica de largo plazo LT-RC (4 semanas) contrarresta la mayoría de estos efectos de expresión de genes produciendo un aplazamiento del envejecimiento. Aunque todavía no se conocen muchas de las respuestas génicas a las diferentes dietas en los humanos, la utilización de microplataformas de ADN es una herramienta poderosa para entender más detalladamente la expresión global de genes celulares en diferentes condiciones nutricionales como en el caso de la presencia o ausencia de oligoelementos en la dieta77-80. Lo más importante es que la genómica nutricional permitirá cruzar la información genómica individual con la alimentación y los componentes de los alimentos, de modo que el efecto sea muy positivo para la salud del individuo. La idea es que los alimentos riesgosos, como las grasas saturadas, podrían reemplazarse con otros potencialmente menos nocivos, que provoquen la descomposición del colesterol. Por ejemplo, comer salvado de avena en lugar de rodajas de queso. No se necesitan millones de dólares para llegar a esta conclusión, pero la ciencia de la nutrición todavía ignora cómo reacciona el organismo en el nivel molecular. La genómica nutricional será la encargada de descubrirlo. El perfil genómico individual puede ayudar a mejorar la nutrición y la salud. La genómica y la bioinformática tienen un gran potencial para identificar genes que causen enfermedades, lo cual se realiza mediante investigación de las bases de datos del genoma humano. Desde esta óptica, los alimentos condicionan la patogénesis de muchas de las enfermedades de la actualidad. En este sentido es fundamental conocer los cambios que produce dentro de la célula su interacción con el transcriptoma y el metaboloma, para poder personalizar los efectos de la dieta sana en la corrección del metabolismo alterado (Gráfica 3). La comparación de un genotipo individual con una base de datos genómica permitirá la prescripción de medicamentos individualizados genotipo dependiente. El mismo enfoque se puede utilizar para el estudio de los metabolitos humanos, con lo cual, en un futuro no muy lejano, se pueden encontrar soluciones personalizadas para mejorar la salud nutricional además de permitir una planificación racional de la producción de alimentos para las siguientes generaciones. Avances en la química analítica han incrementado el conocimiento sobre la estructura de muchos metabolitos que han permitido la consolidación de bases de datos que hacen posible la investigación del potencial de cada uno de ellos. Este progreso ha condicionado un salto gigantesco en la forma como se enfoca la nutrición moderna; sin embargo, todavía es muy temprano para emitir un concepto sobre su impacto en la salud pública. La bioinformática se perfila como una de las herramientas más poderosas en la era postgenómica. Su aplicación en la genómica nutricional debe incluir tres áreas fundamentales:
El futuro de la genómica nutricional. Si bien en un comienzo el cruce entre la dieta y el ADN se hará de manera amplia, la genómica permitirá afinar las prescripciones para una atención preventiva de la salud. Los recientes desarrollos de la proteómica aplicados a la nutrición, están revolucionando los conceptos de alimentos nutriactivos como inductores de la expresión y procesamiento de proteínas cuya acción es fundamental para el funcionamiento normal del metabolismo celular81-83. Probablemente lleve muchos años, pero se considera que la genómica nutricional permitirá organizar dietas que eviten o retarden la aparición de enfermedades graves y muy difundidas hoy, como el cáncer y algunas enfermedades degenerativas como el mal de Alzheimer. Se prevé, además, una dieta que tenga en cuenta a la vez la constitución genética y la actividad. P.e., la dieta de un atleta debe tener en cuenta también su constitución genética para lograr una eficacia máxima84. Ya se han dado algunos pasos para comercializar la investigación en genómica nutricional85. Una empresa denominada Galileo Laboratories ha iniciado conversaciones con fabricantes de alimentos para llevar sus productos al mercado. Galileo se especializa en los problemas de óxido-reducción, perturbación del metabolismo energético celular que provoca enfermedades como la apoplejía, los ataques de corazón, las inflamaciones y la diabetes. La empresa dice que ha encontrado diversos compuestos que podrían prevenir o demorar la aparición de estas enfermedades. La genómica nutricional desempeñará un papel importante para la seguridad porque permitirá comprobar en el laboratorio el efecto de nuevos componentes nutricionales86-88. Pensemos por ejemplo en los ácidos grasos saturados de los alimentos: hace 10 ó 15 años las empresas productoras de alimentos decían que no eran un problema y no tenían efectos negativos sobre la salud y el bienestar. Ahora, sus riesgos son evidentes y la Asociación Cardiológica Norteamericana ha publicado un artículo donde se solicita que los productos alimenticios que los contienen lleven una etiqueta especial, porque pueden provocar el engrosamiento de las arterias89,90. La genómica nutricional es un tema muy complejo que implica preguntas de otro orden que habrá que resolver. El tema de los seguros es una de estas preguntas. ¿Qué consecuencias tendrá para la póliza de seguro de vida el hecho de que el solicitante tenga una tendencia genética a las enfermedades cardiovasculares? ¿Qué consecuencias tendrá para el interesado el hecho de no observar una dieta que demore la aparición de la enfermedad? Todos estos temas todavía imprecisos llegarán algún día a los tribunales. Lo único seguro es que, en el futuro, la dieta tendrá más vinculación con la constitución genética de cada uno y que la fruta diaria se verá reemplazada por algún aminoácido. REFERENCIAS 1. International Human Genome Sequencing Consortium. Initial sequencing and analysis of the human genome. Nature 2001; 409: 860-921. 2. International SNP Working Group. A map of human genome sequence variation containing 1.42 million single nucleotide polymorphisms. Nature 2001; 409: 928-933. 3. World Cancer Research Fund and American Institute for Cancer Research. Food, nutrition and the prevention of cancer: a global perspective. Menasha: BANTA Book Group; 1997. 4. Doll R, Peto R. The causes of cancer: quantitative estimates of avoidable risks of cancer in the United States today. J Natl Cancer Inst 1981; 66: 1191-1308. 5. Venter C, Adams MD, Meyers EW. The sequence of the human genome. Science 2001; 291: 1304–1351. 6. Mathew C. Postgenomic technologies: hunting the genes for common disorders. BMJ 2001; 322: 1031-1034. 7. Nguyen TV, Blangero J, Eisman JA. Genetic epidemiological approaches to the search for osteoporosis genes. J Bone Miner Res 2000; 15: 392-401. 8. Ye SQ, Kwiterovich PO. Influence of genetic polymorphisms on responsiveness to dietary fat and cholesterol. Am J Clin Nutr 2000; 72 Suppl: 1275-1284. 9. Kasim-Karakas SE, Almario RU, Mueller WM, Peerson J. Changes in plasma lipoproteins during low-fat, high-carbohydrate diets: effects of energy intake. Am J Clin Nutr 2000; 71: 1439-1447. 10. Liu S, Willett WC, Stampfer MJ, et al. A prospective study of dietary glycemic load, carbohydrate intake, and risk of coronary heart disease in US women. Am J Clin Nutr 2000; 71: 1455-1461. 11. Eckhardt RB. Genetic research and nutritional individuality. J Nutr 2001; 131 Suppl: 336-339. 12. Emmett A. The human genome. Armed with a working draft, scientist, ponder the meaning of all 23 human chromosome pairs and more. Scientist 2000; 14: 1-3 13. DeRisi JL. Exploring the metabolic and genetic control of gene expression of a genomic scale. Science 1997; 278: 680-686. 14. Searls DB. Bioinformatics tools for whole genomes. Annu Rev Genomics Hum Genet 2000; 1: 251-279. 15. Hacia JG. Resequencing and mutational analysis using oligonucleotide microarrays. Nat Genet 1999; 21 Suppl: 42-47. 16. Fiehn O, Kopka J, Dormann P, Altmann T, Trethewey R, Willmitzer L. Metabolite profiling for plant functional genomics. Nat Biotechnol 2000; 18: 1157-1161. 17. Glassbrook N, Beecher C, Ryals J. Metabolic profiling on the right path. Nat Biotechnol 2000; 18: 1142-1143. 18. Block G, Patterson B, Subar A. Fruit, vegetables and cancer prevention: a review of the epidemiological evidence. Nutr Cancer 1992; 18: 1-29. 19. Peterson J, Dwyer J. Flavonoids: dietary occurrence and biochemical activity. Nutr Res 1998; 18: 1995-2018. 20. Albanes D, Heinonen OP, Huttunen JK, et al. Effects of alpha-tocopherol and beta-carotene supplements on cancer incidence in the alpha-tocopherol beta-carotene cancer prevention study. Am J Clin Nutr 1995; 62 Suppl: 1427-1430. 21. Omenn GS, Goodman GE, Thornquist MD, et al. Risk factors for lung cancer and for intervention effects in CARET, the beta-carotene and retinol efficacy trial. J Natl Cancer Inst 1996; 88: 1550-1559. 20. Risch NJ. Searching for genetic determinants in the new millennium. Nature 2000; 405: 847-856. 21. Mathew C. Postgenomic technologies: hunting the genes for common disorders. BMJ 2001; 322: 1031-1034. 22. Dongre AR, Opiteck G, Cosand W L, Hefta SA. Proteomics in the post-genome era. Biopolymers 2001; 60: 206-211. 23. Guengerich FP. Functional genomics and proteomics applied to the study of nutritional metabolism. Nutr Rev 2001; 59: 259-263. 24. Muller M, Kersten S. Nutrigenomics: goals and strategies. Nat Rev Genet 2003; 4: 315-322. 25. García-Vallejo F. ¿Cómo descifrar el genoma? Innovación y Ciencia 2001; IX: 42-51. 26. Grishin NV, Wolf YI, Koonin EV. From complete genomes to measures of substitution rate variability within and between proteins. Genome Res 2000; 10: 991-1000. 27. Kelloff GJ, Crowell JA, Steele VE, et al. Progress in cancer chemoprevention: development of diet-related chemopreventive agents. J Nutr 2000; 130 Suppl: 467-471. 28. Gohil K. Genomic responses to herbal extracts: lessons from in vitro and in vivo studies with an extract of Ginkgo biloba. Biochem Pharmacol 2002; 64: 913-917. 29. Archer MC, Clarkson TW, Strain JJ. Genetic aspects of nutrition and toxicology: report of a workshop. J Am Coll Nutr 2001; 20: 119-128. 30. Sanderson IR, Naik S. Dietary regulation of intestinal gene expression. Annu Rev Nutr 2000; 20: 311-338. 31. Roberts MA, Mutch DM, German JB. Genomics: food and nutrition. Curr Opin Biotechnol 2001; 12: 516-522. 32. Rock CL, Lampe JW, Patterson RE. Nutrition, genetics, and risks of cancer. Annu Rev Public Health 2000; 21: 47-64. 33. Milner JA, McDonald SS, Anderson DE, Greenwald P. Molecular targets for nutrients involved with cancer prevention. Nutr Cancer 2001; 41: 1-16. 34. Luke A, Cooper R S, Prewitt T E, Adeyemo AA, Forrester TE. Nutritional consequences of the African diaspora. Annu Rev Nutr 2001; 21: 47-71. 35. Peña M, Bacallao J. Malnutrition and poverty. Annu Rev Nutr 2002; 22: 241-253. 36. Wray-Cahen CD, Kerr DE, Evock-Clover CM, Steele NC. Redefining body composition: nutrients, hormones, and Ggenes in meat production. Annu Rev Nutr 1998; 18: 63-92. 37. Cook C, Gatchair-Rose A, Herminghuysen D, et al. Individual differences in the macronutrient preference profile of outbred rats: implications for nutritional, metabolic, and pharmacologic studies. Life Sci 1994; 55: 1463-1470. 38. Sinha R, Norat T. Meat cooking and cancer risk. IARC Sci Publ 2002; 156: 181-186. 39. Martin FL, Ames JM. Formation of Strecker aldehydes and pyrazines in a fried potato model system. J Agric Food Chem 2001; 49: 3885-3892. 40. Friedman M. Prevention of adverse effects of food browning. Adv Exp Med Biol 1991; 289: 171-215. 41. Ames JM, Guy RC, Kipping GJ. Effect of pH and temperature on the formation of volatile compounds in cysteine/reducing sugar/starch mixtures during extrusion cooking. J Agric Food Chem 2001; 49: 1885-1894. 42. Tehrani KA, Kersiene M, Adams A, Venskutonis R, De Kimpe N. Thermal degradation studies of glucose/glycine melanoidins. J Agric Food Chem 2002; 50: 4062-4068. 43. Qiao M, Fletcher DL, Smith DP, Northcutt JK. Effects of raw broiler breast meat color variation on marination and cooked meat quality. Poult Sci 2002; 81: 276-280. 44. Berry BW, Bigner-George ME. Postcooking temperature changes in beef patties. J Food Prot 2001; 64: 1405-1411. 45. Williams RJ. Biochemical individuality. New York: John Wiley & Sons; 1956. 46. Eckhardt RB. Genetic Research and Nutritional Individuality. J Nutr 2001; 131 Suppl: 336-339. 47. Cook C, Gatchair-Rose A, Herminghuysen D, Nair R, Prasad A, Mizuma H, Prasad C. Individual differences in the macronutrient preference profile of outbred rats: implications for nutritional, metabolic, and pharmacologic studies. Life Sci 1994; 55: 1463-1470. 48. Elliott R, Ong TJ. Nutritional genomics. BMJ 2002; 324: 1438-1442. 49.van Ommen B, Stierum R. Nutrigenomics: exploiting systems biology in the nutrition and health arena. Curr Opin Biotechnol 2002; 13: 517-521. 50. Kaplan J. Genomics and medicine: hopes and challenges. Gene Ther 2002; 9: 658-661. 51. Jeffreys AJ, MacLeod A, Tamaki K, Neil DL, Monckton DG. Minisatellite repeat coding as a digital approach to ADN typing. Nature 1991; 354: 204-209. 52. Jeffreys AJ, Wilson V, Thein SL. Hypervariable ‘minisatellite’ regions in human DNA. Nature 1985; 314: 67-73. 53. Armour JA, Crosier M, Malcolm S, Chan JC, Jeffreys AJ. Human minisatellite loci composed of interspersed GGA-GGT triplet repeats. Proc R Soc Lond B Biol Sci 1995; 261: 345-349. 54. Armour JA, Neumann R, Gobert S, Jeffreys AJ. Isolation of human simple repeat loci by hybridization selection. Hum Mol Genet 1994; 3: 599-605. 55. Zane L, Bargelloni L, Patarnello T. Strategies for microsatellite isolation: a review. Mol Ecol 2002; 11: 1-16. 56. Vincent S, Planells R, Defoort C, et al. Genetic polymorphisms and lipoprotein responses to diets. Proc Nutr Soc 2002; 61: 427-434. 57. Schlotterer C. Evolutionary dynamics of microsatellite DNA. Chromosoma 2000; 109: 365-371. 58. Naidoo R, Chetty R. The application of microsatellites in molecular pathology. Pathol Oncol Res 1998; 4: 310-315. 59.Coote T, Bruford MW. Human microsatellites applicable for analysis of genetic variation in apes and Old World monkeys. J Hered 1996; 87: 406-410. 60. Shastry BS. SNP alleles in human disease and evolution. J Hum Genet 2002; 47: 561-566. 61. Lercher MJ, Hurst LD. Human SNP variability and mutation rate are higher in regions of high recombination. Trends Genet 2002; 18: 337-340. 62. Vignal A, Milan D, SanCristobal M, Eggen A. A review on SNP and other types of molecular markers and their use in animal genetics. Genet Sel Evol 2002; 34: 275-305. 63. Judson R, Salisbury B, Schneider J, Windemuth A, Stephens JC. How many SNPs does a genome-wide haplotype map require? Pharmacogenomics 2002; 3: 379-391. 64. Kwok PY. Methods for genotyping single nucleotide polymorphisms. Annu Rev Genomics Hum Genet 2001; 2: 235-258. 65. Nowotny P, Kwon JM, Goate AM. SNP analysis to dissect human traits. Curr Opin Neurobiol 2001; 11: 637-641. 66. Lai E. Application of SNP technologies in medicine: lessons learned and future challenges. Genome Res 2001; 11: 927-929. 67. Brookes AJ. The essence of SNPs. Gene 1999; 234: 177-186. 68. Pietrangelo A. Physiology of iron transport and the hemochromatosis gene. Am J Gastrointest Liver Physiol 2001; 282: G403-G414. 69. Stewart TL, Ralston SH. Review of genetic factors in the pathogenesis of osteoporosis. J Endocrinol 2000; 166: 235-245. 70. Ye SQ, Kwiterovich PO. Influence of genetic polymorphisms on responsiveness to dietary fat and cholesterol. Am J Clin Nutr 2000; 72 Suppl: 1275-1284. 71. Howell WM, Turner SJ, Hourihane JO, Dean TP, Warner JO. HLA class II DRB1, DQB1 and DPB1 genotypic associations with peanut allergy: evidence from a family-based and case-control study. Clin Exp Allergy 1998; 28: 156-162. 72. Nguyen TV, Blangero J, Eisman JA. Genetic epidemiological approaches to the search for osteoporosis genes. J Bone Miner Res 2000; 15: 392-401. 73. Baudin B. Angiotensin I-converting enzyme gene polymorphism and drug response. Clin Chem Lab Med 2000; 38: 853-856. 74. Heller MJ. ADN microarray technology: devices, systems, and applications. Annu Rev Biomed Eng 2002; 4: 129-153. 75. Barzilai N, Shuldiner AR. Searching for human longevity genes: the future history of gerontology in the post-genomic era. J Gerontol A Biol Sci Med Sci 2001; 56: M83-87. 76. Shelley X, Cao, J M, Dhahbi, Mote P L, Spindler SR. Genomic profiling of short- and long-term caloric restriction effects in the liver of aging mice. Proc Natl Acad Sci USA 2000; 98: 10630-10635. 77. Eide DJ. Functional genomics and metal metabolism. GenomeBiology.com 2000; 12: 1028.1-1028.3. 78. Moore JB, Blanchard RK, McCormack WT, Cousins RJ. cDNA array analysis identifies thymic LCK as upregulated in moderate murine zinc deficiency before T-lymphocyte population changes. J Nutr 2001; 131: 3189-3196. 79. Blanchard RK, Moore JB, Green CL, Cousins RJ. Modulation of intestinal gene expression by dietary zinc status: effectiveness of cDNA arrays for expression profiling of a single nutrient deficiency. Proc Natl Acad Sci USA 2001; 98: 13507-13513. 80. Liuzzi JP, Blanchard RK, Cousins RJ. Differential regulation of zinc transporter 1, 2, and 4 mRNA expression by dietary zinc in rats. J Nutr 2001; 131: 46-52. 81. Guengerich FP. Functional genomics and proteomics applied to the study of nutritional metabolism. Nutr Rev 2001; 59: 259-263. 82. Kvasnicka F. Proteomics: general strategies and application to nutritionally relevant proteins. J Chromatogr B Analyt Technol Biomed Life Sci 2003; 787: 77-89. 83. Guengerich FP. Functional genomics and proteomics applied to the study of nutritional metabolism. Nutr Rev 2001; 59: 259-263. 84. Bray MS. Genomics, genes, and environmental interaction: the role of exercise. J Appl Physiol 2000; 88: 788-796. 85. Daniel H. Genomics and proteomics: importance for the future of nutrition research. Br J Nutr 2002; 87 Suppl 2: 305-311. 86. Watkins SM, German JB. Toward the implementation of metabolomic assessments of human health and nutrition. Curr Opin Biotechnol 2002; 13: 512-516. 87. Watkins SM, Hammock BD, Newman JW, German JB. Individual metabolism should guide agriculture toward foods for improved health and nutrition. Am J Clin Nutr 2001; 74: 283-286. 88. Dequin S. The potential of genetic engineering for improving brewing, wine-making and baking yeasts. Appl Microbiol Biotechnol 2001; 56: 577-588. 89. Wang Q. Proteomic approach to coronary atherosclerosis shows ferritin light chain as a significant marker: evidence consistent with iron hypothesis in atherosclerosis. Physiol Genomics 2003; 13: 25-30. 90. Glueck SB, Sigmund CD. Meeting report: physiological genomics of cardiovascular disease: from technology to physiology. Physiol Genomics 2002; 9: 135-136. Copyright 2004 - Revista Colombia Médica The following images related to this document are available:Photo images[rc04025t3.jpg] [rc04025g3.jpg] [rc04025g1.jpg] [rc04025t2.jpg] [rc04025t1.jpg] [rc04025g2.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}