 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Revista Colombia Médica, Vol. 35, Num. 4, 2004, pp. 215-223 Principales factores en el transporte núcleo-citoplasma:
papel de Rev en el proceso del transporte de transcritos de VIH-1
Jorge Eduardo Forero, Bact.1, Silvio Urcuqui, M.Sc., Ph.D.2 1. Estudiante de Maestría en Ciencias Básicas Biomédicas. Grupo de Inmunovirología, Corporación BIOGÉNESIS, Universidad de Antioquia, Medellín. e-mail: jforero@virologia.udea.edu.co 2. Profesor Auxiliar, Laboratorio de Inmunovirología, Facultad de Medicina, Universidad de Antioquia. Grupo de Inmunovirología, Corporación BIOGÉNESIS, Medellín. e-mail: surcuqui@virologia. udea.edu.co usilvio@hotmail.com Recibido para publicación septiembre 30,
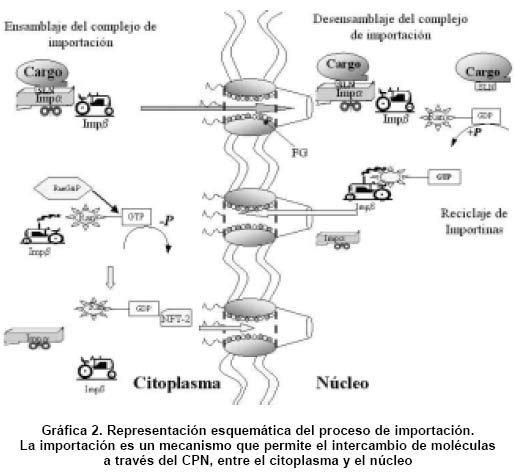
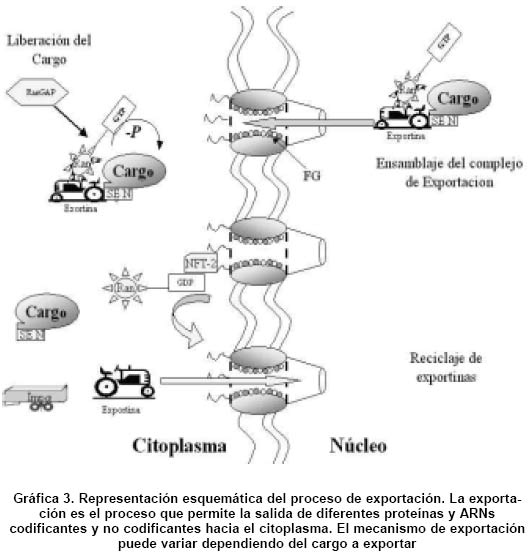
2003 Code Number: rc04043 RESUMEN Las células eucarióticas han desarrollado diferentes mecanismos para la exportación de diferentes ARNs celulares codificantes (ARNm) y no-codificantes (rARN, tARN, snARN), desde el núcleo hacia el citoplasma, donde participan activamente en la producción de proteínas. Las diferentes clases de ARNs son exportadas a través del complejo del poro nuclear (CPN) e intervienen proteínas de la familia de las carioferinas, que pueden ser dependientes o independientes de la proteína Ran. La exportación es un mecanismo específico, coordinado y altamente regulado a través de interacciones proteína-proteína o proteína-ARN. Sin embargo, muchos virus tales como el virus de la inmunodeficiencia humana tipo 1 (VIH-1), han desarrollado un mecanismo de exportación selectivo del ARNm viral. Para tal propósito, el genoma del virus codifica por la proteína Rev, responsable de regular la expresión del genoma viral y de la exportación de los diferentes transcritos virales, interactuando en cis con un ARN altamente estructurado presente en los transcritos virales semiprocesados y sin procesar, en asocio con las proteínas celulares implicadas en la exportación. Con esta revisión se pretende hacer un análisis de los diferentes eventos que ocurren en este proceso, en especial las interacciones proteína-proteína o proteína-ácidos nucleicos. De igual manera, se discutirá la importancia de la proteína Rev del VIH-1, en la exportación de los transcritos virales sin procesar y sus interacciones con las proteínas celulares para llevar a cabo su misión. Palabras clave: Exportación nuclear; Ran; Carioferinas; RRE; Crm1; Poro nuclear. SUMMARY The eukaryotic cells has been developed specific mechanisms for the export of differents classes of coding RNAs (mRNAs) and non-coding RNAs (rRNA, tRNAs and snRNAs) from the nucleus to the cytoplasm, where participates in the process of protein synthesis. The differents classes of RNAs are exported through of the nuclear pore complex (NPC), and are involved protein members of the karyopherin family which could be Ran-dependent or Ran-independent. The export pathway is a mechanism specific, coordinated and higly regulated through to the protein-protein interactions and RNAs binding proteins. However, several viruses such as the human immunodeficiency virus type 1 (HIV-1) have develop a selective mechanism of nuclear export of viral mRNAs. To achieve this goal, the viruses genome code for the Rev protein responsable to regulate the viral genome expression and the export of differents transcripts, by a selective interaction in cis with a hight structure RNA present in the singly spliced and unspliced viral transcripts and interacting with cellular proteins involved in export pathway. This review aims at analyzing the various events that occur during the export process, especially protein (protein and protein) nucleic acid interactions. It also discusses the importance of the HIV-1 Rev protein in the export of the viral unspliced transcripts and the interactions of this protein with the cellular proteins required to fulfill this mission. Key words: Nuclear export; Ran; Karyopherins; RRE; Crm1; Nuclear pore. Las células eucarióticas están organizadas en dos grandes compartimientos de composición y funciones diferentes: el núcleo y el citoplasma. En el núcleo se llevan a cabo procesos esenciales como la trascripción y la replicación, los cuales requieren de factores protéicos y no protéicos que los activen y regulen; en el citoplasma se sintetizan las proteínas indispensables para los procesos nucleares, razón por la cual la célula está equipada con una maquinaria de exportación que permite la circulación de moléculas entre los dos compartimientos de manera rápida, específica y muy regulada1-3. En esta revisión se hace un análisis de los componentes esenciales de la maquinaria de transporte núcleo-citoplasmático celular, haciendo un énfasis muy especial en el mecanismo de exportación de ARNm semiprocesados y sin procesar del VIH-1 y de las proteínas celulares y virales implicadas en este proceso. Complejo del poro nuclear. Con la excepción de un corto período donde la envoltura nuclear pierde continuidad en el proceso de mitosis, todas las macromoléculas transportadas bidireccionalmente entre el núcleo y el citoplasma, deben atravesar el complejo del poro nuclear (CPN). Estos canales especializados permiten la difusión pasiva de iones con diámetros de ~9 nm o proteínas de hasta 40 kDa4,5. Las moléculas que sobrepasen este límite de difusión, son selectivamente transportadas en forma activa (dependiente de energía), por medio de un proceso en el que participan proteínas especializadas, capaces incluso, de translocar moléculas de hasta ~39 nm5. Diferentes estudios1,2 han permitido demostrar que el CPN permite o bien la entrada de las proteínas nucleares o la salida de los diferentes tipos de ARNs. El número de CPNs por célula depende del tipo de organismo, tamaño y actividad biosintética y proliferativa de la célula. En promedio, un núcleo de una célula de mamífero contiene entre 3,000 y 5,000 CPNs6. En los vertebrados, el CPN tiene una masa de ~125 MDa y está compuesto de ~50 polipéptidos distintos que conforman una arquitectura estructural muy conservada entre especies7. Mediante la técnica de microscopía electrónica, se ha logrado establecer que la estructura del CPN es el resultado de la fusión de las membranas interna y externa del núcleo, asociada con otras proteínas que se ensamblan y forman un cilindro asimétrico con respecto a la envoltura nuclear. Ambas caras (citoplasmática y nuclear) presentan un anillo y la parte central está conformada por ocho pliegues proteicos centrales de simetría rotacional; del anillo citoplásmico se desprenden 8 filamentos citoplásmicos, lo mismo que del anillo nuclear, los cuales se proyectan hacia el núcleo y constituyen la estructura conocida como canasta nuclear8-10 (Gráfica 1). El centro del poro está constituido por proteínas que al parecer actúan como transportador. Los componentes individuales del CPN son proteínas conocidas como nucleoporinas y en el caso de la levadura son codificadas por aproximadamente 20 genes. Estas proteínas se caracterizan por la presencia de dominios constituidos por repeticiones múltiples de pequeñas secuencias de fenilalanina-glicina (FG), los cuales se ha sugerido funcionan como sitio de acoplamiento para los factores de transporte núcleo-citoplasmático1,10,11. El ensamblaje de las diferentes nucleoporinas que forman el CPN, parece estar determinado por la nucleoporina Nup107, que hace parte de un complejo hetero-oligomérico formado por Nup160, Nup133, Nup96 y Sec13p. Estudios realizados con Nup107 indican que su depleción, causa una disminución de las otras proteínas del complejo12. Algunas proteínas miembros de la familia de las carioferinas, también juegan un papel muy importante en el ensamblaje de las nucleoporinas. P.e., la carioferina de levadura Kap 121p, participa en el ensamblaje de Nup53p dentro del CPN, gracias a una señal de localización nuclear (SLN) presente en esta nucleoporina13. En el CPN, Nup53p existe en dos complejos separados, uno interactuando con Kap121p y el otro unido a Nup170; la transición entre estos dos estados determinan la liberación o unión de Kap121p con la Nup53p, lo que estaría facilitando el movimiento de Kap121p a través de CPN. Se ha descrito que algunos virus para optimizar la producción de una nueva progenie viral, desarrollan mecanismos que promueven una exportación nuclear selectiva de sus ARNms, que puede interferir con los procesos de transporte celular, e inclusive inhibir dicho proceso. P.e., tanto los rhinovirus como los poliovirus, son capaces de inhibir la importación nuclear mediante la degradación de los componentes del CPN, tales como Nup153 y p6214. Las carioferinas como receptores y adaptadores. Los factores de transporte más estudiados -pero no los únicos- pertenecen a una familia de proteínas muy conservada en eucariontes, conocidas como carioferinas. Estas proteínas son las encargadas de translocar otras moléculas o “cargo”, generalmente proteínas y/o ácidos nucleicos, desde el compartimiento citoplasmático al nuclear y viceversa, respectivamente. Las carioferinas también conocidas como importinas/exportinas, actúan como receptores de cargo y comparten entre sí cuatro características estructurales y funcionales principales: generalmente son de ~90 a ~130 kDa, tienen una región N-terminal que actúa como dominio de unión a Ran-GTP, una región C-terminal de unión al cargo y la capacidad de interactuar con las proteínas que constituyen el CPN. El primer receptor identificado fue la importina b (Imp-b) o carioferina b1, que junto con la importina a (imp-a) constituyen el mecanismo de importación mejor estudiado1-3,15. La Imp-a es un adaptador compuesto de una región N-terminal rica en aminoácidos básicos, conocida como IbB (del inglés Importin b Binding)15, indispensable para la interacción con Imp-b y un dominio central de unión a la señal del cargo. Otros miembros de esta familia de proteínas son Crm1 o exportina 1, y la exportina t, implicadas en la exportación de diferentes clases de ARNs celulares y transcritos virales desde el núcleo hacia el citoplasma11. No está muy claro el mecanismo mediante el cual el cargo es translocado a través del CPN, sin embargo, se ha sugerido que existe una interacción entre las repeticiones FG de las nucleoporinas y los diferentes receptores de transporte, involucrando el anillo de la fenil alanina y residuos hidrofóbicos de la superficie del receptor16. Señales de localización y exportación nuclear. Como se mencionó antes, las carioferinas actúan como adaptadores para otras carioferinas, o se unen directamente a los cargos mediante señales específicas de importación o exportación. Estas señales usualmente están constituidas por repeticiones de una o dos secuencias cortas de aminoácidos y se conocen como señales de localización nuclear (SLN) y señales de exportación nuclear (SEN). Se han identificado dos clases distintas de SLN: monopartita, identificada por primera vez en el antígeno T del virus del simio 40 (SV 40), contiene una sola secuencia corta rica en aminoácidos básicos (PKKKRK) y la bipartita, comprende dos secuencias de aminoácidos cargados positivamente separadas por 12 a 15 aminoácidos de secuencia variable, siendo la secuencia ...KRPAATKKAG QAKKKKLD..., el ejemplo más representativo descrito en la nucleoplasmina1. Las SEN son pequeños motivos ricos en leucina, que tiene como prototipo o secuencia consenso LX2-3LX2-3LXL o residuos hidrofóbicos relacionados. El número de residuos de leucina en la secuencia son variable; pueden afectar su función los residuos de prolina situados entre estos aminoácidos hidrofóbicos6. La primera SEN se describió por primera vez en la proteína viral Rev, responsable de la exportación de los ARNm virales semiprocesados y sin procesar del VIH-1 y la proteína Rex, del virus linfotrópico humano tipo-1, (HTLV-1). Luego se describió en otras proteínas como PKI, aTFIIIA y Gle1p/Rss1p1. Existe un segundo tipo de SEN identificado en la región C terminal de las ribonucleoproteinas heterogéneas nucleares hnRNP, que sirve como señal combinada de importación y exportación1. Es una secuencia de 38 aminoácidos que no comparte ninguna homología con ninguna SEN rica en leucina como la descrita en Rev/Rex; la descripción de estas señales ha facilitado la búsqueda de proteínas con funciones tanto en el núcleo como en el citoplasma. Numerosas proteínas que poseen SLN o SEN, pueden sufrir cambios conformacionales cuando residuos vecinos son fosforilados, llevando a que la señal se oculte, se exponga o disminuya la velocidad de transporte17. Estas alteraciones pueden ser las responsables de la acumulación de proteínas en un compartimiento celular o comportarse como un mecanismo de control de la expresión génica mediada por señales. La proteína Ran, proteína orientadora del flujo de transporte. Una de las características más importantes de las carioferinas es que su función depende de un cofactor clave, la proteína Ran que se requiere para el transporte de sustratos, promueve el ensamblaje o unión de las carioferinas al cargo y es la responsable de la direccionalidad del tráfico de moléculas gracias a su distribución asimétrica en los compartimientos18. Ran se encuentra en dos estados, uno acoplado a GTP (RanGTP) y el otro asociado con GDP (RanGDP). La conversión entre estas dos formas ocurre lentamente en ausencia de factores accesorios. La fosforilación de RanGDP es catalizada por el factor de intercambio de nucleótidos específico de guanina RanGEF19. La hidrólisis de RanGTP es mediada por la proteína activadora de GTPasas específica de Ran, Ran GAP20,21 junto con las proteínas accesorias RanBP1 y RanBP2. RanGEF está confinada en el núcleo, mientras que RanGAP y las proteínas accesorias están restringidas al citoplasma. Esta distribución asimétrica entre los compartimientos es la responsable de la direccionalidad del tráfico de moléculas. Además se debe tener presente que todos los ARNs no codificantes son dependientes de un miembro de las carioferinas y es dependiente de Ran; por el contrario, la exportación de los ARNms es mediado por un mecanismo de exportación independiente de Ran22; sin embargo, ambos mecanismos no están aún muy claros. Proceso general de importación. Los cuatro elementos mencionados (CPN, carioferinas, SLN/SEN, Ran) hacen parte de la maquinaria de transporte nuclear mediado por carioferinas, proceso que se realiza en diferentes etapas:
La importación de las moléculas desde el citoplasma al núcleo comienza con el reconocimiento por la Imp-b de la SLN presente en el cargo a través de su dominio de unión, lo que permite reclutar el adaptador imp-a1-3,11; este proceso es independiente de RanGDP. Paso seguido, el complejo se desplaza hacia el CPN e interactúa con las repeticiones FG de las nucleoporinas y finalmente el complejo con el cargo es translocado al núcleo23. Una vez en el núcleo, RanGDP unido a la Imp-b es fosforilada por RanGAP, lo que provoca un cambio conformacional en el factor de transporte, permitiendo la liberación del cargo en el núcleo. La Imp-b unida a RanGTP regresa al citoplasma donde RanGAP y sus proteínas accesorias la convierten en RanGDP y nuevamente las carioferinas quedan disponibles para un nuevo ciclo de importación2,3,20 (Gráfica 2). La exportación nuclear. Proceso mediante el cual las proteínas y diferentes especies de ARNs codificantes y no codificantes celulares, o ARNm virales, salen desde el núcleo hacia el citoplasma, donde participan activamente en las síntesis de proteínas. La mayoría de estudios se han orientado a comprender la manera como las diferentes clases de ARNs son translocadas al citoplasma como un complejo riobonucleoprotéico (RNP)1-3,11,22. Entre las principales metodologías utilizadas para estudiar la exportación del ARN, están la hibridación in situ, las técnicas de microscopía, las levaduras con mutaciones condicionales que acumulan ARN en el núcleo, detección con anticuerpos específicos, microinyección en huevos de rana y la transfección de células con genes reporteros. La combinación de técnicas ha permitido identificar y caracterizar factores de transporte y a la vez comprender los mecanismos implicados en el proceso de exportación de las diferentes clases de ARNs al citoplasma22. A diferencia de la importación donde sólo el gradiente de Ran permite la translocación del cargo hacia el núcleo, en la exportación RanGTP tiene una participación directa en el reconocimiento del sustrato a transportar y en la estabilización del complejo heterotrímero exportina-Cargo-RanGTP24,25. Si bien no es clara la forma cómo el complejo de exportación interactúa con el CPN, la hipótesis más aceptada es que una vez ensamblado el compejo, la exportina se une transitoriamente con los dominios ricos en FG de las nucleoporinas y permiten el paso del cargo a través del CPN1,6,7,26. En el citoplasma una vez RanGTP interactúa con la región C-terminal de la exportina, es hidrolizado por RanGAP, lo que provoca el desensamblaje del complejo de exportación y la liberación del cargo en el citoplasma. Las carioferinas y Ran son finalmente reingresados al núcleo, según el mecanismo de importación descrito antes. Para mantener en el núcleo las concentraciones de Ran, la proteína transportadora NFT-2 se une a RanGDP libre, promoviendo su paso a través del CPN (Gráfica 3). Exportación de ARNm de la célula. Si bien la mayoría de los ARNs no codificantes celulares son exportado por carioferinas (especialmente Crm1 y Exp t) de manera dependiente del sistema Ran24,26, el ARNm celular es exportado por un mecanismo diferente que involucra las proteínas conocidas como factores de exportación nuclear (NXF) (siglas en inglés de nuclear export factor)22 e independiente de Ran. El ARNm sintetizado por la ARN polimerasa II, una vez asociado con las hnRNP, se acopla con factores proteicos esenciales para su procesamiento, algunos de los cuales además hacen parte del complejo de exportación. El ejemplo mejor estudiado es la proteína de levadura Yra1, implicada en la eliminación de intrones (splicing) y en la exportación22,27-29. De igual manera se ha descrito que la exportación del ARNm, está ligada con las enzimas responsables del encapuchamiento o capping en el extremo 5’y la poliadenilación en 3’ de los ARNm1,22,28,30. Estos resultados sugieren que existe una conección entre la maquinaria de transcripción, la maduración de los diferentes transcritos y el proceso de exportación donde muchas proteínas participan en dos o más eventos. Algunas proteínas que participan en la exportación también juegan un papel muy importante en el control de la expresión génica, en el control de calidad del ARNm (nosense-mediated decay), estabilización del ARNm e iniciación de la traducción31,32. El mediador clave en la exportación del ARNm es la proteína Tap, un heterodímero que junto con el cofactor NXT, se acoplan con los ARNm poliadenilados permitiendo luego una interacción directa con los residuos FG de las nucleoporinas; este proceso se lleva a cabo de manera independiente de Ran31,32. Tap y su equivalente Mex67 en levaduras, presentan un dominio de unión con ARN, una región rica en leucina y un dominio central de acople a NXT (Mtr2p en levaduras)33,34; además tienen una región asociada con la ubiquitina conocida como región UBA. Estudios recientes realizados en levadura muestran que existe una agrupación en la exportación de los ARNm, dependiendo de la función de la proteína que codifican y en dicho proceso intervienen diferentes factores35. Mediante el uso de la técnica de microarreglos de ADN (DNA microarray), se encontró que las proteínas de levadura Mex67 e Yra1, son capaces de exportar 36% y 20% de los ARNm respectivamente36. Estos resultados sugieren que la exportación del ARNm no se realiza de una manera generalizada como tampoco implica el mismo proceso; por el contrario, es un mecanismo que requiere de cierta especificidad de los factores de transporte capaces de reconocer los ARNm directa o indirectamente. MECANISMO DE EXPORTACIÓN DE TRANSCRITOS DE RETROVIRUS SEMIPROCESADOS Y SIN PROCESAR: VIH-1 El ciclo de replicación del VIH-1. El ciclo de replicación del VIH-1, un retrovirus del género lentivirus, es regulado tanto por proteínas virales como celulares y se puede dividir en dos fases: una fase temprana que comienza con el reconocimiento de la célula blanco por el virión maduro gracias a la interacción de receptores de la membrana celular (CD4+) con la proteína viral gp120; este proceso no es suficiente para la entrada del virus, por lo cual necesita de los correceptores celulares CXCR4 o CCR5, que se unen específicamente con la proteína viral gp4137. Una vez en el interior de la célula, el ARN viral es transcrito en una molécula de ácido desoxiribonucléico (ADN) de doble cadena, gracias a la enzima viral transcriptasa reversa. La partícula de ADN resultante se transporta hacia el núcleo como un complejo de preintegración (CPI), donde participan tres proteínas virales con propiedades cariofílicas: la proteína de matriz, la integrasa y Vpr. Finalmente el genoma viral se integra en el genoma de la célula huésped (provirus) por la integrasa viral. Aunque no está bien establecido si la integrasa posee propiedades cariofílicas, contribuye activamente en el transporte del CPI al núcleo38-40. En la fase tardía el provirus toma el control de la maquinaria celular, regula la expresión de su genoma y sintetiza varios tipos de transcritos o ARNm:
Contrario a las células eucarióticas, los retrovirus han desarrollado una estrategia que promueve la exportación específica de sus ARNm semiprocesados o sin procesar, evitando así su retención y degradación en el núcleo. La comprensión del mecanismo de exportación de los transcritos de VIH-1, permitieron comprender la estrategia utilizada por la misma célula para regular la exportación de las diferentes clases de ARNs codificantes y no codificantes al citoplasma. Para lograr su propósito, el VIH-1 codifica por la proteína reguladora Rev, responsable de modular la exportación de los transcritos semiprocesados y sin procesar11. Rev fue el primer factor identificado de exportación nuclear secuencia-específica. Para llevar a cabo su misión, Rev actúa en cis con el elemento de respuesta a Rev (RRE), presente en el gen env de los transcritos a exportar. Durante los primeros estadíos de la infección, el virus muestra un tropismo por los macrófagos (M-trópicos), no hay formación de sincitios y utiliza como correceptor la molécula CCR5. Una vez establecida una infección productiva, el virus se disemina a diferentes órganos y ocurren cambios fenotípicos en el virus, predominando básicamente las cepas con tropismos por las células T (T-trópicos). A diferencia de los M-trópicos, los T-trópicos inducen la formación de sincitios, utilizan como correceptor el CXCR4 e infectan los linfocitos T CD4+. La aparición de las cepas T-trópicas está asociada con la destrucción progresiva de los CD4+ y con el desarrollo del síndrome de inmunodeficiencia adquirida (SIDA). Implicaciones patogénicas del VIH-1/sida. La infección por VIH-1 se caracteriza por la eliminación progresiva de los linfocitos CD4+ (LT CD4+), asociado con una alteración cualitativa y cuantitativa en otras células comprometidas con el sistema inmune42, estado que se conoce como sida. Esto conduce al paciente a una inmnodeficiencia, que lo predispone a infecciones oportunistas y al desarrollo de neoplasias. En términos generales, la patogénesis de la infección por VIH-1 es muy compleja; es un proceso muy variable que depende tanto de factores virales, genéticos e inmunológicos del huésped. Dependiendo del patrón de progresión de la infección, los pacientes se clasifican como progresores rápidos, progresores típicos o progresores lentos43. Además, existe un grupo de individuos clasificados como individuos expuestos seronegativos, quienes han estado expuestos al virus en múltiples ocasiones, pero no muestran evidencia clínica ni serológica de la infección. Este tipo de comportamiento lleva a sugerir la hipótesis de un mecanismo de protección o de resistencia natural a la infección o a la progresión a sida. La eliminación de los LTCD4+ puede ser consecuencia directa de la infección por el VIH-1, debido a una acción directa del mismo virus o a la eliminación de las células infectadas por el sistema inmunitario44; sin embargo, el porcentaje de LTCD4+ infectados es muy bajo en sangre periférica (1/1,000 células en individuos con alta carga viral). La eliminación de los LTCD4+ consecuencia de la infección por el VIH-1 se produce por diferentes mecanismos: consecuencia de la interacción de la proteína de envoltura viral con el receptor CD4, lo que permite la fusión celular y la lisis de la células; liberación de la nueva progenie viral por gemación, lo que produce la destrucción de la membrana celular; por una respuesta citotóxica específica a las células infectadas o por apoptosis inducida tanto por la proteína Tat o por gp12045,46. Exportación del genoma viral. La mayoría de ARNm celulares transcritos por ARN Pol II, se sintetizan como un pre-ARNm, en los cuales se incluyen las regiones intrónicas, que son eliminadas durante el procesamiento (splicing), antes de ser transportados al citoplasma. Una falla en la maduración o una eliminación parcial de los intrones, origina una retención de los transcritos en el núcleo donde finalmente se degradan. Es decir, las células poseen mecanismos de control muy eficaces que les permite hacer un “control de calidad”, gracias a los cuales sólo los ARNm maduros se exportan al citoplasma, donde son reconocidos por la maquinaria de traducción, para la síntesis de las respectivas proteínas. Esto muestra que las células han desarrollado mecanismos que les permite ser muy eficientes en la regulación de su expresión génica en cada una de sus etapas, con claras ventajas para la célula. Sin embargo, en el caso de los retrovirus complejos como el VIH-1, el control y la regulación en la exportación de los transcritos, representa un gran problema a resolver para que no interfieran con su ciclo replicativo. Como se describió antes, el provirus genera tres tipos de transcritos virales: uno de ~2 kb completamente procesado; otro de ~4 kb semiprocesado y el tercero de ~9 kb sin procesar, los cuales deben ser exportados al citoplasma. El transcrito más pequeño es exportado como ARNm celular y da origen a las proteínas Tat, Rev y Nef, que juegan un papel muy importante en la regulación de la expresión génica; sin embargo, para la exportación de los otros dos transcritos, el virus ha desarrollado una estrategia que le permite evadir el sistema de control implantado por la célula. Para lograr este propósito, el virus se sirve tanto de la proteína Rev, responsable de regular la exportación de dichos transcritos y de una secuencia de ARN altamente estructurada localizada en el gen env de los transcritos a exportar47-49. Funciones de la proteína Rev del VIH-1. Rev es una de las proteínas responsables de la regulación de la replicación del genoma viral. Los primeros trabajos mostraron que en ausencia de Rev, los transcritos virales sin procesar permanecían en el núcleo, con efectos dramáticos en la replicación viral49. Después se evidenció que la exportación nuclear de esos ARNm virales, era posible gracias a una acción combinada de Rev y la estructura conocida como RRE presente en todos los transcritos virales semiprocesados y sin procesar47,50,51. La proteína Rev presenta diferentes dominios, que han sido bien caracterizados por mutagénesis. A nivel de la región N-terminal se encuentra una región rica en arginina, implicada en tres funciones fundamentales en el ciclo replicativo del virus:
En la región C-terminal se encuentra un motivo rico en leucina que funciona como dominio de activación y como SEN, ambos implicados en la función de exportación de transcritos de Rev52,53. La presencia tanto de una SLN como de una SEN, permite que Rev pueda translocar desde el núcleo hacia el citoplasma y viceversa. Actualmente existen evidencias que muestran que motivos ricos en leucina como el descrito en Rev, juegan un papel crítico en la localización apropiada subcelular de muchas proteínas celulares y virales54-56. Una vez identificada Rev como la proteína responsable de la exportación de los ARNm sin procesar, muchos investigadores se interesaron en identificar las proteínas celulares implicadas en este proceso. La primera proteína identificada fue la importina b o Crm1, que interactúa con Rev a través de una SEN -presente en ambas proteínas-, una vez se ha formado el complejo Rev-RRE56-59; es decir, en la exportación de transcritos virales la SEN juega un papel fundamental. Es de anotar que Crm1, hace parte del mecanismo de exportación celular. Aunque se conoce parcialmente el mecanismo de exportación al citoplasma de los transcritos virales, se ha establecido que una vez Crm1 se acopla al complejo Rev-RRE, contribuye en el desplazamiento del complejo hacia el CPN, gracias a la asociación con RanGTP58-60; Crm1 se acopla transitoriamente con las nucleoporinas y permite la translocación al citoplasma. También se ha descrito que en cierto tipo de células murinas como A9 (línea celular de fibroblastos), el transporte de los transcritos de VIH-1 es bloqueado posiblemente por la presencia de factores nucleares aún no identificados. Sin embargo, un análogo de Rev, la proteína viral Rex del virus linfotrópico humano tipo 1 (HTLV-1), es funcional en ese tipo de células. Utilizando una proteína quimérica entre la región SLN (aa 1-79) de Rev y la SEN (aa 79-95) de Rex, se logró restaurar la actividad de Rev en células A961. Estos resultados sugieren que la inhibición de la actividad de Rev en esas células se debe a la presencia de uno o varios factores que posiblemente reconocen e interactúan de manera específica con la región activadora (SEN) de Rev, aún sin describir. En forma general, esos resultados llevan a pensar que cierto tipo de células eucarióticas como A9, presentan genes que codifican por proteínas encargadas de bloquear la evolución de algunas enfermedades virales mediante la inhibición o bloqueo de la actividad de proteínas virales fundamentales en el ciclo de vida del virus e implicadas en la patogenicidad o en la replicación61. Igualmente se han descrito algunas proteínas celulares con funciones homólogas de Rev. Así p.e., se ha demostrado que la proteína celular Sam68 (Src-associated protein in mitosis), es capaz de interactuar con la estructura viral RRE y sustituir o actuar de manera sinérgica con Rev en la exportación nuclear, expresión génica y replicación viral62,63. CONCLUSIONES Y PERSPECTIVAS El transporte de moléculas a través del poro nuclear, es uno de los procesos más sorprendentes de la función celular. Los avances de la ciencia y la tecnología en los últimos años, han permitido resolver preguntas claves sobre el proceso que permanecieron por mucho tiempo sin respuesta. Es así como actualmente se sabe que existe una íntima relación entre los procesos que se realizan en el núcleo y el citoplasma, el cual se ve reflejado en el control de la expresión génica. Así, la interferencia en los eventos de procesamiento de los ARNm provocan una ineficiente translocación de éstos hacia el citoplasma con consecuencias graves en la síntesis de las proteínas. Existe una conexión interesante entre los factores o proteínas responsables del transporte y los componentes del complejo del poro nuclear. En este sentido, a pesar del extraordinario progreso logrado en la comprensión del transporte celular, aún quedan muchas preguntas clave sin respuesta en particular en lo que tiene que ver con la exportación de moléculas (ARNs y proteínas) de origen viral. Un aspecto a recalcar, es el sistema utilizado por los retrovirus más complejos para evadir los mecanismos de retención nuclear de transcritos sin procesar, cuyo producto es necesario para su replicación viral y generación de una nueva progenie. En el caso de VIH-1, la presencia de la proteína Rev y su capacidad de interactuar con los diferentes factores de la maquinaria de transporte, le han permitido al virus desarrollar una eficiente exportación de sus ARNm. La caracterización estructural y funcional de Rev ha ayudado no sólo a entender los aspectos esenciales de la exportación en eucariontes, sino también ha contribuido en la búsqueda de nuevas estrategias de control de la replicación del VIH-1. Una mejor comprensión de las vías de exportación, podría claramente ayudar a identificar nuevos blancos potenciales con fines terapéuticos. En ese sentido, por las características y el papel fundamental que juega la proteína Rev en la regulación de la expresión del material genético del VIH-1, se convierte en un blanco muy atractivo en la búsqueda de nuevas estrategias en la lucha contra la infección y en la búsqueda de un mejor bienestar de las personas con sida. Esto es muy importante si se tiene en cuenta que muchos virus son capaces de desarrollar resistencia a ciertos antiretrovirales. REFERENCIAS
© Copyright 2004 - Revista Colombia Médica The following images related to this document are available:Photo images[rc04043f2.jpg] [rc04043f1.jpg] [rc04043f3.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}