 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
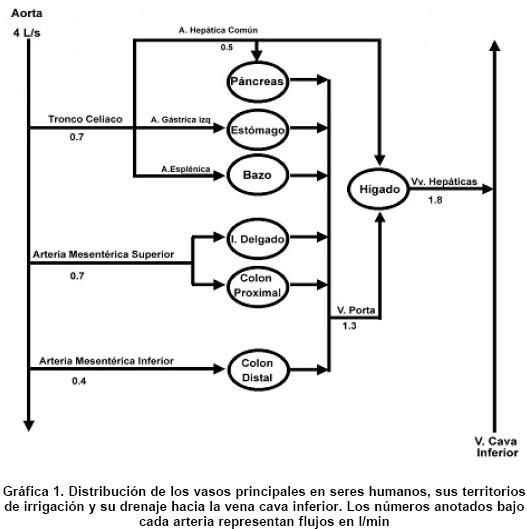
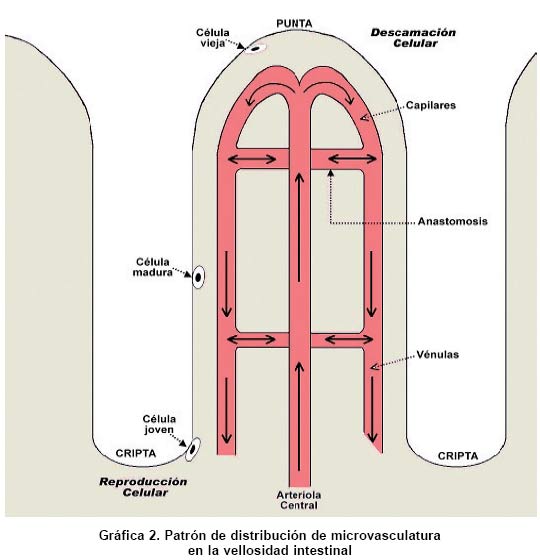
Revista Colombia Médica, Vol. 35, Num. 4, 2004, pp. 231-244 Circulación intestinal: Su organización, control y papel en el paciente crítico Raúl Narváez-Sánchez, M.D., M.Sc.1, Lilian Chuaire, M.Sc.2, Magda Carolina Sánchez, Lic. Quím.3, Javier Bonilla,M.D.4 1. Instructor Asistente, Coordinador Laboratorio
de Fisiología, Instituto de Ciencias Básicas, Facultad de Medicina,
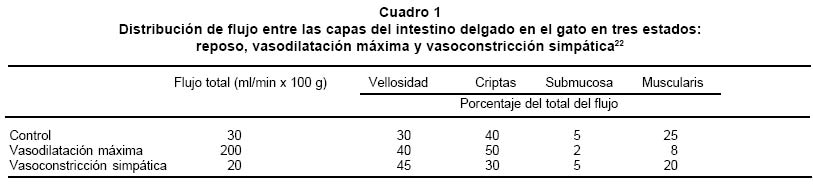
Universidad del Rosario, Bogotá. e-mail: rnarvaez@urosario.edu.co Recibido para publicación mayo 25, 2004 Code Number: rc04045 RESUMEN El intestino ha ganado un espacio fundamental en el campo de la monitorización y del manejo de pacientes en unidades de cuidados intensivos, porque este órgano puede desempeñar tanto un papel protector, como un papel promotor en la insuficiencia multiorgánica del enfermo crítico. Por tal motivo desde hace 20 años, la fisiología intestinal ha dado base a un área de investigación floreciente, en la carrera por validar métodos mínimamente invasivos que permitan efectuar un diagnóstico temprano y una monitorización efectiva del choque (p.e., tonometría, espectrometría de impedancia y oximetría de pulso endoluminal). Con el fin de mejorar la comprensión del funcionamiento y del control de la circulación sanguínea en el tubo digestivo, sobre todo en el estómago y en el intestino del ser humano, se presenta una revisión actualizada que parte de un recuento elemental de la distribución anatómica e histológica comparada de los vasos sanguíneos del abdomen y de la pared intestinal. También se revisa de modo breve la biofísica aplicable. Por último, se estudian los mecanismos reguladores del flujo sanguíneo en el tubo digestivo, con actualizaciones sobre la información clásica. Palabras clave: Fisiología; Circulación intestinal; Insuficiencia multiorgánica. SUMMARY Intestine has gained a fundamental room in monitoring and treatment of the patient in the Intensive Care Unit, since this organ can switch its roll from protector to promoter of multiple organ failure in critical patients. Moreover, this organ is an interesting research field in last 20 years, running to validate minimally invasive methods for early diagnosis and monitoring of shock (for example, tonometry, impedance spectrometry, and endoluminal pulse oxymetry). For a better understanding of function and control of blood circulation in the digestive tube, an updated review about that physiology is presented, with an emphasis in human stomach and intestine. First of all, we present an elemental and comparative inventory of the anatomic and histological distribution of blood vessels in abdomen and in the intestinal wall; then, we review some applicable biophysics; finally, we study the regulatory mechanisms of blood flow in the digestive tube, updating on the classical information. Key words: Physiology; Circulation intestinal; Multiple organ failure. La insuficiencia multiorgánica, también llamada falla multisistémica o falla orgánica múltiple, es la causa principal de muerte en salas de cuidados intensivos. La mucosa intestinal es reconocida como fundamental en el proceso de descompensación y choque, porque la isquemia en ella puede cambiar el papel de la mucosa, de limitador a promotor de la insuficiencia multiorgánica. “La isquemia mesentérica aguda puede resultar de émbolos, trombosis arterial y venosa, o vasoconstricción secundaria a bajo flujo. Las tasas de mortalidad informadas en los últimos 15 años permanecen tan altas como estaban hace más de 70 años y promedian 71%, con un rango de 59% a 93%. El diagnóstico antes del infarto intestinal es el factor más importante para mejorar tan pobres resultados”1. Es determinante para el diagnóstico, guía de terapia y pronóstico, identificar no sólo presencia de isquemia, sino cuantificar el daño ocasionado y la consecuente viabilidad del tejido afectado2. Es necesario posicionar al intestino como órgano central a cuidar en el paciente crítico y para ello se requiere conocer cómo se regula su flujo sanguíneo y por ende cuáles son los posibles puntos de alteración de ese flujo, lo que puede conducir a isquemia intestinal e insuficiencia multiorgánica. El tubo digestivo es un órgano grande (aproximadamente 300 m2 de superficie, donde 250 corresponden al intestino delgado)3, que posee los mecanismos básicos de un ser vivo independiente, incluyendo un sistema nervioso primitivo. La circulación intestinal cumple dos funciones básicas: proveer oxígeno a los tejidos y transferir los nutrientes absorbidos al resto del cuerpo. Esta última tarea la hace junto con la circulación linfática. La gran área del tubo digestivo y de su circulación debe permitir estas dos funciones, pero también tiene que limitar la posible pérdida de proteínas o de cualquier sustancia desde la sangre hacia el exterior. Por otra parte, esa gran área es motivo de activa investigación, pues desde allí se puede monitorizar, o incluso aumentar, el grado de oxigenación tisular del paciente4. La circulación esplácnica es la parte de la circulación sistémica que irriga la porción abdominal del tubo digestivo, así como el bazo, el páncreas y el hígado. Es la circulación regional más grande nacida de la aorta y recibe más de una cuarta parte del gasto ventricular izquierdo durante el reposo, independientemente de que el sujeto esté recién alimentado o no5. Este volumen equivale a unas diez veces el flujo sanguíneo que recibe toda la masa muscular esquelética en reposo, cifra enorme si se tiene en cuenta que esta masa tiene el mayor lecho vascular del cuerpo humano6. Ese gran tamaño determina que la circulación esplácnica sea el más importante reservorio de sangre de todo el cuerpo. Así, cuando la presión sanguínea tienda a disminuir, la vasoconstricción intestinal desplazará cerca de una quinta parte de la volemia hacia los órganos que la requieran. En animales como el caballo, el bazo participa también como reservorio de sangre, mientras que en el ser humano, en apariencia, carece de esta función. Esta revisión sobre control del flujo sanguíneo intestinal se divide en tres partes: recuento anatómico, recuento biofísico y revisión de los eventos mecánicos y moleculares, hasta hoy conocidos, que regulan el flujo sanguíneo intestinal, haciendo énfasis en particularidades del paciente crítico. ANATOMÍA Para los propósitos de esta revisión, la circulación intestinal se dividió en: 1. Vasos principales: tronco celíaco y arterias mesentéricas superior e inferior. 2. Vasos intermedios: arcos que distribuyen la irrigación desde las mesentéricas. 3. Microvasculatura: de trayecto intramural. 4. Venas: distribuidas de forma paralela a las arterias. 5. Vasos linfáticos.Distribución de los vasos principales. Las principales técnicas usadas para estudiar la anatomía de la macrocirculación intestinal son la disección cadavérica y la cateterización arterial selectiva. Tronco celíaco (TC). Se desprende de la aorta (Gráfica 1), a la altura de la primera vértebra lumbar. Su longitud es algo más de un centímetro y su diámetro cercano a la mitad del aórtico. El TC emite tres ramas: la arteria esplénica, que irriga al bazo, la arteria gástrica izquierda, que irriga al cardias y la curvatura menor estomacal y la arteria hepática común, que vierte su contenido en la cabeza pancreática, en la primera parte del duodeno y en el hígado. El territorio irrigado por el TC es también suplido por ramas de las arterias intercostales inferiores provenientes de la pared abdominal, así como por las ramas frénicas de la aorta y por las ramas esofágicas bajas. Por ello, a pesar de su gran tamaño, la oclusión del TC causa una perturbación relativamente pequeña al riego de los órganos abdominales altos. La isquemia causada por oclusión del TC no alcanza a alterar sensiblemente la función de estos órganos7. Arteria mesentérica superior (AMS). Emerge desde la aorta (Gráfica 1), aproximadamente 1 cm por debajo del origen del TC. Es la fuente principal de irrigación intestinal. Su diámetro es similar al del TC. El ángulo que forma su origen y el eje longitudinal de la aorta es menor a 90°, lo que le permite capturar un importante caudal, pero también posibles émbolos. En la AMS se originan ramas muy anastomosadas. Estas son las arterias intestinales, que irrigan a todo el intestino delgado, la arteria cólica media, que irriga al colon transverso, la arteria cólica derecha, que irriga al colon ascendente y la arteria ileocólica, que surte al íleon terminal y al ciego. A diferencia de los territorios irrigados por el TC, los que son surtidos por la AMS tienen muy poca circulación colateral y por ello una oclusión aguda de su tronco principal puede desencadenar la muerte, a veces en pocas horas. De hecho, los estudios de isquemia intestinal inducida se realizan, en casi todos los protocolos, mediante pinzamiento de la AMS sobre el mesenterio. Las vías colaterales que irrigan a los territorios de la AMS son los vasos pancreáticos, las arcadas gástricas y las gastroepiplóicas y los vasos extracelómicos. Si el bloqueo de la AMS es gradual, da lugar a la formación de nuevas vías anastomóticas que son normales en su histología y fisiología8. Arteria mesentérica inferior (AMI). Emerge de la aorta (Gráfica 1) unos centímetros antes de su bifurcación. Irriga la parte final del colon transverso, el descendente, el sigmoide y la parte proximal del recto. El diámetro de la AMI es aproximadamente un tercio del de la AMS y cuenta con una circulación colateral abundante. Distribución de la microvasculatura. Los métodos usados son la inyección y/o corrosión en especímenes preparados, así como la fluorescencia de rayos X. En adición con las técnicas histológicas convencionales, se aplica al tejido un medio de contraste radiográfico como el sulfato de bario y luego se efectúan los procedimientos de fijación, corte y microrradiografía. La microvasculatura intramural Distribución. En la pared del tubo digestivo, desde la luz intestinal hasta el peritoneo, hay cuatro capas o túnicas: la mucosa, la submucosa, la muscular propia y la túnica serosa o adventicia. En el estómago, los vasos submucosos se ramifican en capilares en la base de las glándulas gástricas. Luego pasan en forma perpendicular a través de la mucosa, donde terminan en una red subepitelial de capilares que drena en las vénulas de la lámina propria de la mucosa. Luego estas vénulas convergen hacia las vénulas colectoras que pasan al plexo submucoso. Presenta una arquitectura similar en el intestino delgado y en el grueso. La microcirculación nace en las arterias llamadas vasa recta, que reciben caudal sanguíneo de los vasos intermedios. Las vasa recta primero dan unas ramas que forman el plexo seroso. A continuación penetran, de manera oblicua, en la pared intestinal hasta llegar a la submucosa, donde dan origen a un nuevo plexo, que queda entremezclado con linfáticos y con el plexo ganglionar nervioso submucoso o de Meissner. El plexo vascular submucoso es pues una red tortuosa y muy anastomosada, donde se originan vasos más pequeños que irrigan la mucosa, o recurren hacia la muscular propia. Así, la muscularis propria recibe dicha irrigación recurrente, pero además recibe vasos del plexo seroso, de manera que es muy resistente a la isquemia. En estudios de isquemia en que se pinza la AMS, es habitual encontrar la muscularis propia casi sana cuando ya la túnica mucosa está completamente necrosada. Esta susceptibilidad de la mucosa a la isquemia se debe también a que la circulación que discurre por ella es de tipo terminal y transporta el flujo sanguíneo apenas necesario para ejecutar las funciones de esta capa. Cuando se considera que todas las funciones intestinales se realizan en la mucosa, salvo la actividad motora, es explicable su abundante irrigación, pero también su mayor sensibilidad a la isquemia respecto de las demás capas. La microvasculatura de la mucosa, en contraste con la intramural, presenta diferente arquitectura en el intestino delgado comparada con la del intestino grueso. Así, en el colon, las ramas del plexo submucoso forman un auténtico plexo mucoso basal. Los vasos emanados de este plexo son rectos y paralelos y no se comunican entre sí a lo largo de sus dos tercios proximales. En el tercio distal emiten múltiples ramas que se anastomosan formando un plexo vascular mucoso superficial, que visto desde la luz intestinal, parece un panal de vasos rodeando a las criptas de Lieberkühn. Entre especies, la arquitectura microvascular del intestino delgado varía considerablemente. En el ser humano se encuentra un plexo mucoso basal semejante al del colon, desde donde se originan los vasos que, de manera individual, irrigan a cada vellosidad (Gráfica 2). Este único vaso se llama arteriola central (AC) y pierde su capa muscular lisa de manera paulatina desde la base de la vellosidad hacia su punta. En el ápex de la vellosidad, la AC se arboriza en múltiples capilares que, cual largas agujas de una sombrilla o chorros que se separan en el vértice de una fuente, regresan hacia la base de la vellosidad, donde se anastomosan en una densa red justo por debajo del epitelio. En el humano y el perro, la red subepitelial drena a las vénulas submucosas antes de llegar a la base de la propia vellosidad. En cambio, en el gato esta red sólo drena a dichas vénulas cuando ha alcanzado la base de la vellosidad. La arquitectura descrita acarrea dos consecuencias: la primera es que la microvellosidad tiene cierta autonomía funcional con respecto de los vasos intramurales. La segunda consecuencia es que puede ocurrir un intercambio contracorriente entre la sangre arterial y la venosa, lo que facilita la hipoxia en la punta de la vellosidad. La mayor parte de la experimentación en isquemia intestinal se efectúa en modelos animales fuertes como el perro, el cerdo y la rata. Aunque el conejo tiene la microcirculación intestinal muy similar a la del ser humano, no constituye un buen modelo por su baja resistencia a las condiciones experimentales. FISIOLOGÍA DE LA CIRCULACIÓN INTESTINAL Se puede modelar el intestino como un tubo recubierto externamente, que tiene una capa media donde radica su capacidad de movimiento y una capa interna que hace absorción y secreción. Regulación del flujo en un vaso principal. La ecuación de Poiseuillea evidencia que el principal factor regulador del flujo en un vaso sanguíneo es el radio, pues es el único factor no lineal. Al reordenar esta ecuación, se observa que también es el radio el principal determinante de la resistencia al flujo de sangre, porque la resistencia varía inversamente con la cuarta potencia del radio. a. Flujo= p/8 (N (P1-P2) r4/hL), donde p/8=constante de proporcionalidad; N=Número de tubos paralelos, idénticos en radio y longitud, en que se distribuye el flujo; (P1-P2)=Diferencia de presión entre los extremos del vaso; r = radio del vaso; h=viscosidad de la sangre total; L = longitud del vaso. Como la capa media de los vasos principales es más elástica que muscular, la modificación de su radio es pasiva. Estos son vasos de distribución y no de resistencia. Es decir, “se dejan llenar” y luego fluctúan alrededor de su diámetro inicial, de modo que empujan la sangre hacia delante, sin aumentar la resistenciab,9,10. b Esta propiedad de las arterias grandes se llama elastancia, que matemáticamente se expresa como el inverso de la pendiente dV/dP (razón de cambio del volumen sobre razón de cambio de la presión). Mayor elastancia, menor resistencia. En los vasos principales el diámetro y los esfuerzos son lo suficientemente altos como para que la sangre se comporte como un fluido newtoniano11,12. Esto hace que no se modifique la viscosidad h ni factor alguno de la ecuación de Poiseuille, de modo que se cumple a cabalidad esta ecuación. Por lo señalado, dentro de lo normal los vasos principales no tienen gran importancia en la regulación del flujo sanguíneo a sus territorios irrigados. La presión interna en los vasos principales es virtualmente idéntica a la presión central aórtica. Además, el ángulo de emergencia de estos vasos a partir de la aorta es favorable para no alterar demasiado el régimenc laminar del fluido. Bastaría calcular el número de Reynolds en un segmento de un vaso principal distante de una bifurcación, para encontrar que su flujo es laminar y por tanto veloz y con menor gasto de energía que si el flujo fuera turbulento9. c Organización de la sangre durante su desplazamiento a lo largo de un vaso. Regulación del flujo en la microvasculatura. La microcirculación del tracto gastrointestinal presenta características anatómicas y mecanismos fisiológicos que le permite ajustarse a los requerimientos tisulares complejos propios de este tracto. En particular, este órgano presenta una mayor densidad capilar que la mayoría de los demás órganos (se calcula una longitud capilar vellosa de 100 km/100 g de mucosa, comparada p.e., con los 30 km/100 g de tejido muscular esquelético). Con el fin de analizar el modo en que funciona la microvasculatura, se puede considerar que el lecho vascular intramural está constituido por secciones en serie y por secciones en paralelo (Gráfica 3). Cuando cambia la demanda metabólica de alguna capa, el arreglo en paralelo debe satisfacer esa demanda, ayudado por anastomosis que existen entre una y otra capa. Esto permite que, a pesar de desplazamientos ocasionales de sangre entre capas, no cambie el volumen sanguíneo total requerido por el intestino. En resumen, un arreglo en serie vierte sangre al plexo vascular submucoso y este distribuye a los arreglos en paralelo según la intensidad de trabajo de cada tejido, sea muscular o mucoso. Los elementos en serie siguen en las vénulas y terminan en las venas. Arteriolas y meta-arteriolas. Son vasos precapilares. Se les llama musculares por la abundancia de músculo liso en su capa media. Gracias a que pueden cambiar el tono de su músculo liso y, por tanto, el radio del vaso, las arteriolas y meta-arteriolas son las principales determinantes de la resistencia al flujo sanguíneo hacia una región particular, a la vez que se constituyen en el sitio de control tanto remoto como local para regular la tasa de flujo sanguíneo. Los esfínteres precapilares contribuyen poco a la resistencia vascular total regional, pero sí controlan el número de capilares perfundidos. Así, modifican tanto la distancia media que hay para la difusión entre la sangre y el tejido, como el tiempo disponible para el tránsito capilar. Por tanto, regulan el tiempo disponible para el intercambio de sustancias. El reajuste hemodinámico en las arteriolas acarrea, por una parte, incremento del volumen sanguíneo perfusor, lo que quiere decir mayor número de vellosidades perfundidas y por otra parte, aumenta el tiempo medio de tránsito sanguíneo a lo largo de la vellosidad. Capilares. Son los vasos de intercambio. En el intestino hay capilares tanto continuos como fenestrados. Según la ecuación de Fickd, donde el árbol vascular tenga un área máxima y un mínimo espesor de pared, tienden a intercambiarse sustancias solubles y de poco peso molecular, cuya concentración difiera a ambos lados de la pared del vaso. En cuanto al área capilar, una de las mayores áreas de la circulación en mamíferos corresponde a los capilares intestinales13. En el ser humano se calcula un área capilar de 1.9 m2/100 g de microvellosidades, lo que en total equivale al área de un campo de tenis. En cuanto al espesor, el grosor de la pared capilar es similar al de la barrera aire-sangre, es decir, menos de una millonésima de metro. Esto se debe a que esta pared está constituida sólo por endotelio, que separa los compartimentos intra y extravascular. Los capilares fenestrados aumentan la conductividad hidráulica, lo que permite el intercambio de pequeños solutos (electrolitos y nutrientes) a la vez que restringe la pérdida de componentes macromoleculares (coloidales) desde el plasma. d Flujo=Difusibilidad [área vaso (presión capilar–presión intersticial)]/espesor de la membrana Vénulas. Son vasos de resistencia postcapilar. El tono de esta sección vascular es un determinante de la presión hidrostática media capilar y por ende modifica la tasa de intercambio de fluidos a través de la pared capilar (ecuación de Fick). La constricción de las vénulas incrementa la presión hidrostática capilar, de modo que aumenta la filtración de fluido desde el espacio intravascular hacia el extravascular14. En las vénulas comienza la sección vascular de capacitancia, que se prolonga a lo largo de las venas hasta regresar al corazón. Cuando cambia el tono venular, el volumen sanguíneo regional se altera, pero la resistencia vascular total regional no cambia de modo significativo15,16. Aunque en la submucosa se encuentran shunts arteriovenosos, varios estudios han demostrado que el flujo relativo a través de estos vasos es menos de 1% del flujo total, por lo que parecen tener una importancia fisiológica pequeña17. La gran conductividad hidráulica de los capilares de la mucosa digestiva representa un riesgo para el paciente crítico, debido a que un pequeño incremento en la presión hidrostática capilar podría translocar grandes volúmenes de plasma al espacio extravascular. Por ello existen varios mecanismos que tratan de mantener esta presión a un nivel casi constante, a pesar de notorias variaciones en la presión arterial del individuo. Uno de estos mecanismos, quizá el más importante, es la relación entre la resistencia precapilar y la postcapilar16,18,19. REGULACIÓN DEL FLUJO SANGUÍNEO EN EL INTESTINO Regulación local en el intestino.El primero en medir el flujo de sangre al intestino fue Mall, en 1888, citado por Lundgren20 quien trabajó en perros. A partir de su investigación, se conoció que el flujo sanguíneo gastrointestinal guarda más relación con la intensidad de la función, que con el soporte vital mismo del tracto digestivo. La autorregulación de este flujo (ya sea en términos de flujo propiamente dicho, o en términos de mantener una presión hidrostática constante en el capilar) es menos eficiente en el intestino que en el riñón o en el cerebro, pero más que en el músculo esquelético. Esta autorregulación permite que el flujo sanguíneo general a todas las capas se encuentra casi constante, aun a pesar de variaciones en la presión de perfusión entre 80 y 160 mmHg. La respuesta autorreguladora radica principalmente en las arteriolas precapilares y varía entre los circuitos paralelos. P.e., los vasos en las vellosidades tienen una mayor capacidad autorreguladora que los del resto de las capas intestinales. Por ello el flujo en la parte absortiva del intestino se mantiene prácticamente constante, a pesar de reducciones en la presión de perfusión incluso hasta 30 mmHg21. Esta autorregulación se explica por dos mecanismos. El primero, que se llama miogénico o “mecánico”, consiste en que el músculo liso de un vaso intestinal se contrae al aumentar la presión transmural o viceversa, así como hay contracción precapilar al aumentar la presión venosa. El segundo mecanismo, que se llama “metabólico”, autorregula el flujo mediante relajación del músculo liso secundaria a acumulación de metabolitos vasodilatadores, de modo similar a como sucede con el flujo sanguíneo cerebral cuando se acumulan iones hidrógeno. El intestino recibe más de la mitad de la circulación esplácnica, en proporcionalidad con su área. En muchas especies, incluido el hombre, hay un gradiente de flujo sanguíneo por gramo de tejido (ml/100 g/min), máximo en el duodeno y mínimo en el colon. El intestino delgado recibe cerca de dos veces la sangre que recibe el colon, lo que es consistente con un gradiente correspondiente en el consumo de oxígeno. Del flujo total que el intestino recibe en reposo, 70% va al epitelio (criptas y vellosidades). Al elevarse el flujo, se eleva el volumen que va a la mucosa, hasta alcanzar 90% del total. La vasodilatación en el epitelio roba sangre a las capas más profundas (Cuadro 1). El estímulo más potente para aumentar el flujo sanguíneo intestinal es la ingestión de alimento, de manera que la actividad metabólica intestinal se multiplica para digerir y absorber. La entrada del quimo a la luz intestinal eleva, en un mayor o menor grado, el flujo sanguíneo a la mucosa, según el tipo de constituyentes de ese contenido. P.e., en el yeyuno, el flujo sanguíneo aumenta unas 10 veces más si recibe alimento digerido mezclado con bilis, respecto de si recibe sólo alimento no digerido, sólo bilis o sólo aminoácidos23. La presencia del quimo en la luz intestinal causa, en términos de motilidad, interrupciones en el complejo mioeléctrico migrante (actividad eléctrica intrínseca del músculo liso), de manera que en la fase digestiva la actividad motora presenta un patrón irregular pero efectivo que permite mezclar los contenidos intestinales, a la par que los pone en contacto con la máxima área posible de mucosa. El músculo sin actividad rítmica cede parte de su irrigación24, de manera que durante esta fase la mucosa seguirá teniendo la mayor parte del flujo sanguíneo. Entre sus funciones normales, el intestino debe secretar y absorber. En cuanto a la secreción, se considera que es una forma de transporte activo de solutos en que la filtración transcapilar tiene escasa participación, incluso en estados de severa hipersecreción tales como el cólera. En cuanto a los procesos de absorción, la sangre debe proveer sustratos energéticos para que las células mucosas absorban activamente algunas sustancias. Es difícil determinar hasta qué punto se altera la tasa de absorción de los solutos absorbidos de manera activa cuando cambia sólo el flujo sanguíneo, o si esta alteración se debe a un efecto indirecto del metabolismo celular. Es más fácil medir el grado de la alteración con solutos absorbidos en forma pasiva. En ratas se ha determinado que la tasa de absorción de compuestos lipofílicos se eleva con el aumento del flujo sanguíneo. En cambio, la tasa de absorción de sustancias hidrosolubles, con excepción de la propia agua, no se afecta ni siquiera por grandes variaciones de flujo sanguíneo. Es decir, la velocidad de paso de estas sustancias a través del epitelio intestinal es el punto limitante a su propia tasa de absorción25. El peristaltismo afecta la circulación intestinal de dos maneras opuestas. Por una parte, puede elevar el flujo sanguíneo por acumulación de metabolitos, del mismo modo que el ejercicio acarrea hiperemia en el músculo esquelético. Por otra parte, puede disminuir el flujo al exprimir segmentos que se están contrayendo. Aun así, el movimiento per se no alcanza a elevar significativamente el flujo total intestinal, a pesar de un posible pequeño desvío de sangre hacia la muscularis, desde otras capas24. La mucosa no se afecta por tal desviación de flujo, en parte por su autorregulación y en parte por la rica red de anastomosis que hay en la submucosa. Otros factores metabólicos que regulan las microcirculaciones locales y que se han propuesto como mediadores importantes en la microcirculación gastrointestinal, son la adenosina y la tensión tisular de oxígeno. La adenosina es un potente vasodilatador en el lecho esplácnico y hay evidencia de su acumulación durante las hiperemias reactiva y postprandial en el intestino delgado. Esta vasodilatación se modifica al interferir la acción o el metabolismo de la adenosina, ya sea por cambios en la presión arterial o en la absorción de nutrientes26. El papel de la tensión local de oxígeno en la regulación vascular intestinal es menos claro. Cuando se reduce la liberación de oxígeno o se aumenta su demanda, o ambos, cambia la perfusión de modo proporcional. Sin embargo, no se puede determinar el papel de la tensión tisular de oxígeno per se en esta motricidad vascular, debido a la acumulación concomitante de otros metabolitos vasodilatadores. Por otra parte, a un determinado consumo de oxígeno, la magnitud de la hiperemia postprandial es mayor si ha sido alta la extracción de oxígeno en reposo (determinada por la diferencia arteriovenosa de O2)27. Hay otros posibles reguladores locales de la circulación intestinal, como prostaglandinas, angiotensina, histamina, serotonina, hiperosmolalidad, iones potasio, purinas y purinonucleótidos. De estas, se ha observado que las prostaglandinas y la angiotensina regulan la motilidad vascular del tracto gastrointestinal, pues actúan como factores homeostáticos en la microcirculación. La histamina es un potente vasodilatador, pero no se le ha encontrado un papel fisiológico en el intestino. Quizá en una situación patológica, sobre todo donde haya mastocitosis en la mucosa o la serosa, la histamina pueda desempeñar un papel importante26,28. Un ejemplo de situación patológica es la colitis ulcerativa fulminante. En pacientes críticos aquejados por esta enfermedad, se han medido flujos sanguíneos colónicos de hasta 150 y 200 ml/min/100 g, cuando el valor normal en reposo es menor de 30 ml/min/100 g. Esto implica que el flujo total al colon puede alcanzar 1500 ml/min, es decir, 30% del gasto cardíaco en reposo. Además del posible papel de la histamina, evidencia reciente soporta que en la enfermedad inflamatoria del intestino, las plaquetas expresan niveles significativamente mayores del ligando CD40, que induce la producción de proinflamatorios en el endotelio microvascular29 y una consecuente y severa vasodilatación. Regulación hormonal del flujo sanguíneo en el intestino. En la actualidad se considera que el intestino es la glándula más grande del cuerpo humano. Aparte de las hormonas ya nombradas, otras hormonas tienen efectos hemodinámicos experimentales en el intestino, pero sólo cuando son aplicadas en concentraciones superiores a aquellas consideradas fisiológicas (p.e., concentraciones encontradas después de comer). Las más investigadas de estas hormonas son la colecistoquinina (CCK), gastrina, polipéptido gástrico inhibitorio (GIP), glucagón, neurotensina y secretina. Muchas de las hormonas gastrointestinales liberadas al comer son vasodilatadoras cuando se inyectan directamente en la circulación esplácnica, pero ello no constituye evidencia de que esas sustancias puedan vasodilatar a las concentraciones que se lograrían fisiológicamente durante o después de la ingesta. De las hormonas citadas , en particular la CCK, secretina, gastrina y neurotensina, si se inyectan solas o combinadas en una arteria a dosis que se alcanzarían en el período postprandial, no logran producir vasodilatación. En cuanto al glucagón, éste estimula el flujo sanguíneo intestinal, en una respuesta que, al menos en cerdos, es mediada por el óxido nítrico30,31. Por su parte, el péptido intestinal vasoactivo (VIP) sí logra producir vasodilatación a concentraciones postprandiales o “fisiológicas”. Además, el bloqueo del VIP endógeno suprime la hiperemia inducida por infusión intraluminal de bilis y ácido maléico. Así, el VIP liberado por las neuronas del plexo submucoso parece ser fundamental en la hiperemia intestinal. El VIP es el único péptido en el intestino que actúa por la vía del AMPc, tan sólo compartida por la toxina del cólera. El VIP produce importante relajación del músculo liso arteriolar y consecuente vasodilatación. El mecanismo específico de esta relajación muscular ocurre, al menos en parte, sobre canales de potasio sensibles al ATP [K(ATP)], presentes en el músculo liso vascular, que también responden a una variedad de vasodilatadores farmacológicos y endógenos. Los canales K(ATP) se componen de cuatro subunidades de rectificación entrantee que sirven de canal para el K+ y cuatro receptores de sulfonilurea, que actúan como reguladores. Los canales K(ATP) son inhibidos por ATP intracelular y por agentes sulfonilúricos. Ciertos vasodilatadores farmacológicos como el cromakalim, el pinacidil y el diazóxido activan directamente los canales K(ATP). La consecuente hiperpolarización membranal cierra los canales de Ca2+, voltaje-dependientes, produciendo reducción del Ca2+ intracelular, relajación del músculo liso y vasodilatación. Los vasodilatadores endógenos VIP, péptido relacionado con el gen de la calcitonina (CGRP)26, prostaciclina y adenosina, activan los canales K(ATP) por estímulo a la formación de AMPc e incremento de actividad de la proteincinasa A. Al menos una parte del mecanismo de contracción inducida por los vasoconstrictores endógenos se debe a la inhibición de los canales K(ATP). Estos canales K(ATP) son también inhibidos bajo condiciones patológicas como hipoxia, isquemia, acidosis y choque séptico. En consecuencia, estos canales juegan un papel clave para la regulación de la perfusión tisular32, sobre todo en el paciente de cuidados intensivos. e Canales de rectificación entrante: el término entrante de “inward”, indica que estos canales tienden a mantener el estado membranal de despolarización; el término rectificador se debe a que su relación de corriente respecto al voltaje -la conductancia- no es lineal. La corriente es no rectificada u óhmica cuando la resistencia de la membrana es constante en un rango de potenciales membranales. Cuando la conductancia varía con el potencial de membrana, es decir, la resistencia membranal depende del voltaje, se habla de que tiene canales rectificadores. En el paciente crítico es posible encontrar los eventos patológicos arriba nombrados, debidos p.e., a hemorragia severa. En este caso la respuesta vasoconstrictora es mediada en parte por hormonas. Las que tienen más efectos demostrados en este tipo de injuria son las catecolaminas circulatorias, la angiotensina y la vasopresina33-35. La respuesta neuroendocrina inducida por estrés mayor, produce aumento de hormonas contrarreguladoras tipo catecolaminas, cortisol y glucagón. Estas hormonas, en gran parte mediante inducción de una relativa resistencia a la insulina, buscan aumentar los sustratos energéticos disponibles, p.e., a través glucogenolisis, gluconeogénesis, inhibición de síntesis proteica, y movilización de grasas desde los adipocitos. Así, en el paciente crítico hay dos características opuestas: hiperglicemia y resistencia a la insulina. Se inhibe la cetosis, que ahorraría proteínas y toda la respuesta catabólica produce una rápida reducción de la masa magra corporal y discapacidad para reponer hormonas proteicas y enzimas. Por ello una preocupación fundamental del intensivista debe ser la reposición agresiva de nutrientes, sea por vía enteral o parenteral. El comportamiento de la circulación intestinal en el paciente crítico que recibe nutrición enteral, depende completamente de su balance cardiovascular e hidroelectrolítico, y en particular de su capacidad de oxigenar el tejido, habida cuenta de que la absorción de sustancias en el intestino consume una importante cantidad de energía. Se suele recomendar el inicio temprano de la vía enteral (a más tardar 48 horas luego de la admisión a UCI), para reducir la posibilidad de recurrir a la vía parenteral. El clínico debe tener en mente que por el catabolismo, el paciente crítico tiene menos secreción de moco y éste es pobre en IgA. Las uniones intercelulares en la vellosidad intestinal están débiles, de modo que usar la vía parenteral como única fuente de nutrientes puede comprometer aun más la función de barrera y favorecer la traslocación bacteriana. Esta misma debilidad del intestino explica que en algunas personas el inicio demasiado temprano de la vía enteral produce mayor morbimortalidad. Es importante anotar que la nutrición parenteral en períodos cortos, comparada con nutrición exclusivamente enteral, no altera las concentraciones séricas de gastrina, colecistocinina, VIP, neurotensina ni bombesina36. En ese orden de ideas, el uso alternativo de nutrición parenteral por períodos cortos de hasta dos días, no causaría alteraciones significativas en el control vasoactivo intestinal. Por otra parte, mediciones realizadas en cerdos recién nacidos muestran que la transición de nutrición enteral a parenteral puede inducir en menos de 8 horas disminución en el flujo sanguíneo intestinal, lo que tiende a aumentar la atrofia vellosa. Esta transición produce también supresión de la síntesis proteica en 24 horas y suspensión de la proliferación celular en criptas, amenazando la supervivencia a 48 horas37. Regulación neural del flujo sanguíneo en el intestino. El flujo sanguíneo se eleva a partir de la fase cefálica de la digestión, como cuando se ve o se saborea la comida38. Durante la ingesta el flujo sanguíneo intestinal prácticamente se duplica. Incluso, de los resultados obtenidos por diversos autores23,39 puede inferirse que es posible, que el acto de imaginar la comida tenga un efecto similar en el aumento del flujo sanguíneo al intestino. Esto desde la fase cefálica es mediado por el sistema nervioso central, en especial por el hipotálamo y el tallo cerebral, con aumento de la actividad vagal. Gracias a los avances en la medición de actividad arteriolar intestinal, hoy se conoce que la vasodilatación neurogénica en las arteriolas submucosas se debe a la acción de la acetilcolina y/o otros neuropéptidos liberados por neuronas intrínsecas submucosas, así como a la liberación de sustancia P y CGRP desde nervios extrínsecos sensitivos. Estas dos vías vasodilatadoras se caracterizan, primero porque pueden activarse de modo independiente por la estimulación mucosa provocada por el alimento y segundo, porque ambas tienen componentes aferentes y eferentes en los plexos neuronales mucoso y submucoso. Vanner et al.39 plantean que las vías entéricas intrínsecas, colinérgicas reflejas, están implicadas en el control local fisiológico de flujo sanguíneo a la mucosa, mientras que las vías extrínsecas, sensitivas reflejas, se activarían de manera preferente en estados no fisiológicos. Control simpático. En 1855, Pflüger demostró que el intestino tiene abundantes fibras simpáticas adrenérgicas, que inducen vasoconstricción cuando se estimulan eléctricamente. Por fuera de la pared intestinal, las arterias mesentéricas presentan abundante inervación simpática. Dentro de la pared, hay inervación adrenérgica cerca de los vasos de la muscularis, la submucosa y en las criptas de la mucosa (Gráficas 2 y 3). La inervación vasoconstrictora en las arteriolas de la capa submucosa está mediada sólo por nervios simpáticos extrínsecos que liberan ATP en receptores purínicos P2n arteriolares39. En las vellosidades hay poca inervación simpática y poca musculatura lisa, de manera que en los vasos de la vellosidad no puede radicar una importante vasomotricidad activa, como sí sucede en las criptas. Dresel et al.40 estudiaron en gatos los efectos de las fibras vasoconstrictoras en secciones vasculares consecutivas, con un método pletismográfico combinado con difusión transcapilar de Rb86. Al iniciar la estimulación de las fibras constrictoras, se encontró respuesta de las arteriolas precapilares, con muy pobre respuesta de vasos de capacitancia y de esfínteres precapilares. De manera sorprendente, tanto el flujo como el volumen y la presión retornan a sus valores iniciales apenas transcurridos dos a cuatro minutos después del inicio de la estimulación y sin que ésta se haya suspendido, fenómeno que se llama “escape”. Esto significa que, de alguna manera, los vasos sanguíneos intestinales dejan de obedecer la vasoconstricción inducida por el simpático, como una respuesta autorreguladora. El escape podría deberse a que el músculo liso hace una adaptación de sus receptores a-adrenérgicos, o bien, que haya activación concomitante de receptores ß-adrenérgicos, que causan vasodilatación41,42. Desde un punto de vista funcional, el patrón de respuesta descrito implica que la tasa de absorción de solutos no es afectada por el sistema nervioso simpático, aunque la secreción y la tasa de mitosis en las criptas podrían decrecer si el flujo sanguíneo cae por debajo de un nivel crítico. Hay sitios del sistema nervioso central cuya estimulación modifica la influencia vasoconstrictora regional sobre el tracto gastrointestinal. P.e., estimular el hipotálamo anterior produce una reacción de defensa (o alarma) que incluye el ataque o la huida y una importante vasoconstricción de todo el intestino. La estimulación simpática también puede reflejarse al reducir la presión en el seno carotídeo y/o al aumentar la actividad quimiorreceptora. Esta actividad quimiorreceptora puede ser inducida a su vez, por aumento de la PaCO2, por disminución de la PaO2, o por disminución del pH arterial, concomitantes en cualquier combinación, o en forma separada. Se observa una respuesta similar en animales cuando las aferencias somáticas de presión son estimuladas en los nervios periféricos. Estas fibras probablemente conducen estímulos dolorosos desde la superficie del cuerpo. El dolor visceral, en contraste, evoca una caída en la presión arterial e incremento en el flujo sanguíneo, como resultado de inhibición de la actividad simpática43. Control parasimpático. Debido a la abundante inervación parasimpática en todos los órganos abdominales, por largo tiempo se creyó que el intestino también la tenía. Pero hasta la fecha, con las mejores técnicas disponibles, no se han encontrado fibras parasimpáticas vasodilatadoras en el intestino delgado. Pero la estimulación eléctrica vagal produce vasodilatación en el intestino, del mismo modo que sucede en las coronarias. Ello nos permite concluir que la citada vasodilatación en la fase cefálica de la digestión obedece a factores metabólicos y/o hormonas locales, liberados por estímulo parasimpático en el epitelio y no por acción directa. En esta respuesta al estímulo eléctrico vagal se ha identificado al VIP como el principal mediador44-46. En el colon no se logra elevar el flujo sanguíneo al estimular el vago, aunque sí al estimular los nervios pélvicos. Es decir, aunque la influencia del parasimpático sobre el tubo digestivo puede elevarse fisiológicamente en varias condiciones, en particular eventos emocionales, al parecer sus consecuencias vasodilatadoras no son ejercidas de manera directa en ninguna parte del intestino. En resumen, aparentemente el principal mecanismo de regulación neural del flujo a la microcirculación intestinal es la variación del tono vasoconstrictor simpático. El patrón de flujo en la mucosa es ante todo afectado por la influencia adrenérgica nerviosa, aunque no induce una marcada redistribución del flujo en las capas de la pared intestinal. A pesar de que no hay fibras parasimpáticas vasodilatadoras en la submucosa, se han identificado reflejos intrínsecos dilatadores en los vasos de esta capa, que involucran fibras colinérgicas. Estas fibras al parecer actúan sobre el receptor muscarínico endotelial M3, que a su vez activa la liberación de óxido nítrico, potente relajante del músculo liso arteriolar. En estos procesos participan varios neuropéptidos además del VIP, como son la sustancia P y la CGRP. Es interesante resaltar que la sustancia P parece contribuir particularmente en la regulación de la circulación intestinal del recién nacido, en especial durante los primeros días posteriores al nacimiento47,48. INTERCAMBIO CONTRACORRIENTE INTESTINAL La expresión “intercambio contracorriente” significa el desplazamiento de sustancias desde un espacio donde se mueve fluido en un sentido, hacia otro espacio donde se mueve fluido en el sentido contrario. Según lo descrito en la distribución microvascular dentro de la vellosidad intestinal, se desprende que es factible el intercambio contracorriente en ésta, pues la dirección total del flujo capilar subepitelial se opone a la dirección de la sangre en el centro de la vellosidad (Gráfica 2). La distancia entre estos flujos contracorriente es cercana a 20 mm, teniendo en cuenta que la sección transversal de la vellosidad puede ser cilíndrica (como en el gato) o elipsoide (como en el conejo o la rata). Además de la distancia entre los espacios vasculares, hay otros factores fundamentales para el intercambio contracorriente, como son la permeabilidad y el tiempo de tránsitof. Como el volumen vascular de la red capilar subepitelial es aproximadamente nueve veces mayor que el de la arteriola central, el tiempo de tránsito en esa red será 9/10 de las velocidades de tránsito anotadas. Se ha medido la velocidad de tránsito de eritrocitos en la vellosidad intestinal en gatos49 y en ratas, y se ha encontrado en “reposo” una velocidad de 0.5 mm/s en los capilares. Esta velocidad se reduce en el sujeto con choque hemorrágico, pero lo hace significativamente más en personas con endotoxemia50. f Tiempo de tránsito: Tiempo que tarda un fluido, en este caso la sangre, en pasar por una sección de tubería. P.e., en la microvasculatura pulmonar es de 0.75 segundos, tiempo en el que un eritrocito pasa cerca de unos tres alvéolos. El tiempo de tránsito es determinante para el intercambio que alcanza a hacer la sangre con los tejidos. La primera implicación funcional del intercambiador velloso intestinal es que una parte de los solutos absorbidos desde la luz intestinal se mueve en pro del gradiente de concentración, hasta la arteriola central en su parte no muscular. Además, desde la arteriola central se moverán solutos hacia la red capilar subepitelial, en mayor cantidad cuanto más próximos se esté a la base de la vellosidad. Es muy importante resaltar que el oxígeno es una de las sustancias que se moviliza desde la arteriola central hacia la red capilar subepitelial, de modo que se crea un gradiente de tensión de oxígeno desde la base de la vellosidad hacia su punta. Esto hace crítica la cantidad del gas que llega a las células en el ápex de la vellosidad. La tensión de oxígeno en la punta de la vellosidad es casi cero51,52. Este cortocircuito (shunt) es reforzado por el alto consumo de oxígeno que hacen las células más próximas a la base vellosa. Las células epiteliales de la vellosidad se multiplican en las criptas (Gráfica 2), es decir, en la base y van envejeciendo a medida que llegan hacia el ápex. Por ello y por la hipoxia a la que se las somete en tal proceso, es obvio que sean las células de la punta las que más descamen, para ser reemplazadas desde la base. La segunda implicación funcional del intercambiador velloso intestinal consiste en que este es un multiplicador de solutos hidrosolubles y transportados “activamente”, como el sodio. El sodio se transporta hacia los capilares en una solución levemente hipertónica, por lo que puede difundir en los sentidos ya descritos. Sin embargo, la permeabilidad al sodio es probablemente baja en la pared capilar que da hacia la arteriola central, pues las fenestraciones están en la cara que da hacia las células epiteliales. Por otra parte, la célula endotelial capilar que da hacia la arteriola central probablemente tiene mayor permeabilidad al agua que al sodio. Así, la absorción activa de sodio produce una diferencia de presión osmótica entre los dos espacios del intercambiador y fuerza al agua a pasar desde la arteriola central hacia los capilares. Por la disposición que tienen los vasos en la punta de la vellosidad, la pequeña diferencia en la concentración de sodio entre los capilares y la arteriola se multiplica a lo largo de la vellosidad, de modo similar a como sucede en las nefronas. Por ello, en el ápex velloso la osmolalidad es de 700 a 800 mOsm/Kg H2O en la punta, lo que crea la fuerza suficiente para absorber agua desde la luz intestinal. Así, el agua se absorbe exclusivamente por difusión osmótica simple, mientras que los iones inorgánicos se absorben por transporte activo. Sólo los iones cloruro pasan por difusión pasiva53. El vaso linfático llamado quilífero central, es un tubo que va desde la punta hacia la base de cada vellosidad. Una de sus funciones es absorber los quilomicronesg con diámetro menor a 0.5 mm. El tamaño de los quilomicrones les impide atravesar la pared de los capilares sanguíneos vellositarios y por eso los linfáticos son la principal vía de absorción de estos y otros lípidos hacia la sangre. Las vellosidades tienen movimiento activo mediante contracción de su músculo liso longitudinal. Con una frecuencia aproximada de seis veces por minuto, cada contracción acorta la vellosidad hasta 50% de su longitud, lo que reduce su capacidad, de modo que impulsa hacia la base el contenido del vaso quilífero central y los capilares sanguíneos. Estos movimientos responden además a la estimulación mecánica, a la presencia de quimo y a la hormona villicinina. g Quilomicrones: Partículas grasas de triglicéridos, colesterol y fosfolípidos unidos con proteínas. REGULACIÓN DEL FLUJO SANGUÍNEO EN EL ESTÓMAGO En el estómago, la distribución de la sangre es similar a la del intestino. En el estómago, prima el mecanismo metabólico sobre el mecánico, porque allí el flujo sanguíneo parece depender mucho menos del movimiento que de la actividad secretora. Se estima que en reposo, 70% del flujo sanguíneo estomacal se distribuye hacia la mucosa. Como en el intestino, este valor se eleva cuando hay mayores tasas de flujo. A tasas secretoras pico, cerca de 90% del flujo total gástrico perfunde la mucosa54. La secreción gástrica, bien sea estimulada con histamina, gastrina o acetilcolina, o bien sea inhibida con catecolaminas, serotonina55 atropina y/o secretina, induce una variación del flujo hacia la mucosa de manera proporcional. Otras sustancias que incrementan el flujo sanguíneo a la mucosa gástrica son la gastrina, secretina y pancreocimina. Por otro lado, la somatostatina disminuye ese flujo56-58. Diversos autores han trabajado sobre los efectos que tiene la motilina sobre el flujo sanguíneo en la mucosa gástrica56,59. A ese respecto, Jin et al.60 demostraron recientemente que el flujo sanguíneo gástrico de perros se eleva en fase con las contracciones digestivas y la secreción estomacal. Según sus hallazgos, la motilina puede inducir este aumento sostenido del flujo en las arterias gástricas, sin afectar variables como la presión sanguínea general, la frecuencia cardíaca, o el flujo en la arteria mesentérica superior. Las respuestas a la motilina, como son las contracciones gástricas, la secreción y los cambios fásicos del flujo en la arteria gástrica izquierda, fueron abolidas mediante la aplicación de atropina o del antagonista de motilina GM-109 (fe-ciclo[Lis-Tir(3-tBu)-betaAla]trifluoroacetato). También observaron que la atropina, a diferencia del GM-109, no tiene mayor efecto en el incremento del flujo sanguíneo, a menos que se hayan usado dosis pequeñas de motilina. Las respuestas a motilina en la arteria gástrica izquierda no son inhibidas por el granisetron (antagonista de la serotonina), ni por bloqueadores H2, ni por bloqueadores a y ß-adrenérgicos. Otro hallazgo interesante de Jin et al. consistió en que la motilina induce vasodilatación significativamente mayor que la inducida por dosis equivalentes de VIP. En resumen, la motilina es un vasodilatador gástrico potente y selectivo, mediado en su efecto probablemente tanto por mecanismos colinérgicos como no colinérgicos. El mecanismo autorregulador metabólico tiene a su vez, dos mecanismos que explican la estrecha relación secreción-flujo sanguíneo. Uno de estos mecanismos es indirecto y el otro, directo. El indirecto es la actividad metabólica celular, si se tiene en cuenta que el consumo de oxígeno por parte del estómago aumenta proporcionalmente con la secreción de ácido. Esto indicaría que los estímulos o las inhibiciones a la secreción modifican indirectamente el flujo, por liberación de vasodilatadores en el tejido. Este mecanismo es dependiente del nivel de flujo sanguíneo, porque a altas tasas de flujo (que proveen adecuado volumen de oxígeno y remueven bien los metabolitos secretados por la mucosa) está casi ausente. El mecanismo directo es mediado por la actividad vasomotora directa de los secretagogos o de los inhibidores de secreción gástrica. P.e., la acetilcolina y la histamina son vasodilatadores gástricos, mientras que la norepinefrina y la vasopresina son vasoconstrictores. Debido a que este mecanismo directo es independiente del flujo, los factores que sólo modifican el flujo por la vía indirecta lo harán menos, por unidad de secreción, que aquellos con acción vasomotora directa. Es importante aclarar que un vasodilatador directo aumentará el flujo, pero no hasta el punto en que el flujo sanguíneo deje de ser un factor limitante para el incremento en la secreción. Como el nivel de secreción excede el aumento en el flujo sanguíneo causado por la vasodilatación directa, el incremento extranecesario será aportado por los factores metabólicos locales, de modo que se mantiene un cambio similar en el flujo sanguíneo, con respecto al cambio en la secreción. En sujetos sanos fumadores y no fumadores, se ha comprobado que la nicotina reduce el volumen de secreciones estomacales, la secreción de ácido y el flujo sanguíneo a la mucosa, de manera dependiente de la dosis. Sonnenberg61 observó que el volumen de secreciones está más inhibido que el flujo sanguíneo a la mucosa y por tanto concluyó que la nicotina eleva el flujo sanguíneo a la mucosa, relativo a la secreción gástrica reducida. No encontró asociación de la nicotina con el desarrollo de úlcera péptica y propuso que su posible acción ulcerogénica se debe llevar a cabo por mecanismos diferentes al cambio en la secreción ácida y en el flujo sanguíneo a la mucosa. En el paciente crítico, es muy frecuente el desarrollo de úlcera péptica. La evidencia sobre el papel de la microcirculación gástrica en su fisiopatología muestra que la lesión mucosa precede y no sucede, al compromiso microcirculatorio. Es conocido que la restauración de la circulación ayuda a proteger la mucosa de los agentes ulcerantes, del mismo modo que la insuficiencia circulatoria contribuye a aumentar el tamaño del daño mucoso. Esto se explica cuando se considera la mucosa gastrointestinal como una interfaz entre los contenidos luminales -sean nutrientes o sustancias peligrosas- y las estructuras submucosas. El ácido gástrico es uno de los principales componentes injuriosos del contenido luminal. Para protegerse, el epitelio tiene un sistema de “alerta temprana” que activa, entre otras defensas, la regulación del pH intracelular (pHi) epitelial, el flujo sanguíneo y la secreción de gel mucoso. A concentraciones fisiológicas de ácido luminal, las células epiteliales disminuyen su pHi, el flujo sanguíneo se eleva (respuesta hiperémica) y aumenta el espesor de la capa de moco. La respuesta hiperémica al ácido puede abolirse mediante el bloqueo del intercambio Na+/H+, los receptores vanilloides (VR)h, los receptores al CGRP y la sintasa del óxido nítrico (NO), o por denervación sensorial aferente. Sin embargo, este efecto no se logra cuando se usa un inhibidor no selectivo de la ciclooxigenasa (COX). La secreción de moco como respuesta al ácido puede disminuir al interrumpir la vía de la capsaicina, lo que incluye bloquear el VR, los nervios aferentes sensibles a capsaicina, CGRP y NO y puede abolirse mediante inhibición de la COX. Esto ha permitido postular que la vía de la capsaicina es sensible al ácido y, en respuesta a la acidificación luminal, promueve la hiperemia y la secreción de moco. La vía de la COX ocupa un lugar secundario para regular la secreción de moco. Existe también en el colon una vía de capsaicina, sensible al ácido, similar a la gástrica. Es como si la mucosa gastrointestinal “saboreara” la acidez luminal mediante una comunicación epitelio–receptor vanilloide62. CONCLUSIONES El conocimiento detallado de los mecanismos de regulación del flujo sanguíneo intestinal es un campo activo de investigación y publicación mundial. Para realizar un manejo razonable del paciente en estado crítico, el médico necesita conocer a fondo tanto la fisiología intestinal, como la fisiopatología circulatoria del tubo digestivo. Este órgano, cuando se enferma, pierde su capacidad de barrera para las bacterias, y pasa a ser un promotor de la falla multisistémica. Es indispensable que en cada unidad de cuidado intensivo se disponga de algoritmos de reposición nutricional basados en una carta metabólica del paciente, para un inicio de la vía enteral temprano y agresivo, pero razonado. AGRADECIMIENTOS Sincero agradecimiento al cirujano Andrés Isaza y al neurofisiólogo Carlos Moreno, por sus valiosos comentarios a esta revisión. También a la señora Nancy Cantor, por la realización técnica de las gráficas. REFERENCIAS
© Copyright 2004 - Revista Colombia Médica The following images related to this document are available:Photo images[rc04045t1.jpg] [rc04045f3.jpg] [rc04045f2.jpg] [rc04045f1.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}