 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
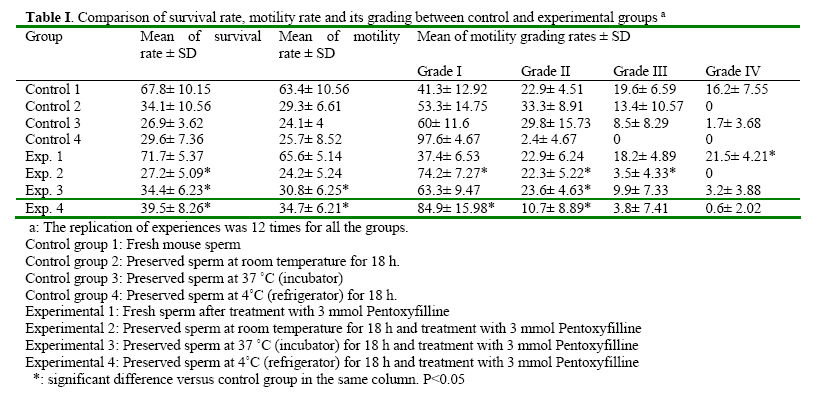
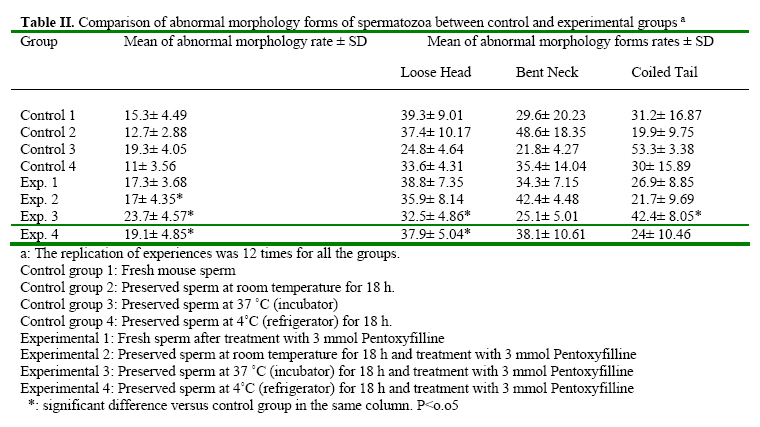
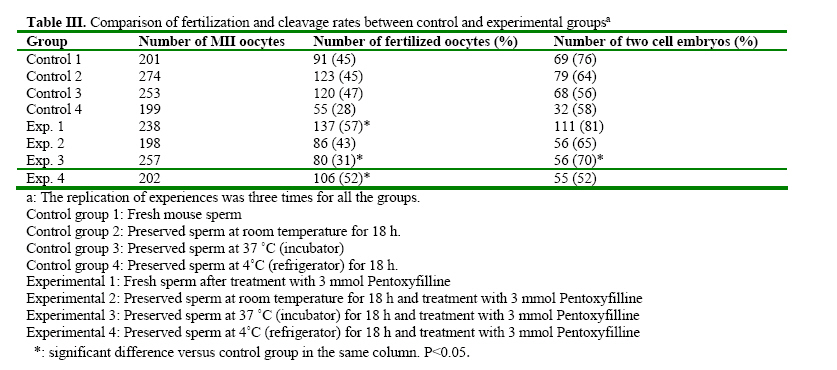
Iranian Journal of Reproductive Medicine, Vol. 2, No. 2, 2004, pp. 51-57 The Effects of Pentoxifylline on Mouse Epididymal Sperm Parameters, Fertilization and Cleavage Rates after Short Time Preservation Iraj Rashidi, M.Sc,, Mansoureh Movahedin, Ph.D., and Taki Tiraihi, Ph.D. Department of Anatomy, Medical Sciences Faculty, Tarbiat Modarres University, Tehran, Iran. Code Number: rm04011 Background: Pentoxifylline (PX) prevents cAMP breakdown by inhibiting the activity of the cAMP-phosphatase and presumably, stimulates sperm motion. Incubation with PX causes hyperactivation of sperm, an important step in achieving fertilization, and leads to changes in membranes associated with sperm capacitation. Key words: Pentoxifylline, Mouse sperm, Motility, Short time preservation Introduction Cryopreservation is widely used in many assisted conception units to preserve male fertility, for example before cytotoxic chemotherapy, radiotherapy (Sanger et al, 1992), or certain surgical treatments that may lead to testicular failure or ejaculatory dysfunction. Unfortunately, the use of cryopreserved spermatozoa limits the success of these techniques because the freezethaw process results in reduced motility, viability and fertilization potential of human semen (Crister et al., 1987). Freezing and thawing involve damage to the plasma membrane and acrosome of human spermatozoa as evidenced by significant ultra structural changes demonstrated by electron microscopy (Wooly and Richardson, 1987). Despite many refinements in cryopreservation methodology, the salvage of post thaw sperm remains poor (Agrawal et al., 1995). Contrary to belief, the challenge to cells during cryopreservation is not their ability to endure storage at low temperature; it is the lethality of an intermediate zone of temperature (-15 to~0°C) that cells must traverse twice, once during cooling and once during warming (Mazur, 1963). As a result, some investigators tend to preserve sperm at the conditions other than -196 ºC, for example preservation at 4 ºC for short time is recommended (Fuller and Whittingh, 1996; Nassar et al., 1998; Tao et al., 1995). Many substances have been proposed for stimulation of human sperm function in vitro. These include poorly defined biologic materials, (e.g., serum, peritoneal fluid, and follicular fluid) as well as defined agents, such as adenosine analogues, progesterone and methylxanthines (e.g., caffeine and pentoxifylline) (Tesarik et al., 1992; Tournay et al., 1995). Pentoxifylline (PX) is a methylxanthine that acts as a phosphodisterase inhibitor and subsequently increases intracellular cyclic adenosine monophosphate (cAMP) levels. Since the reports by Yovich et al. (1988 & 1990), methylxanthines, particularly PX, again have become popular in assisted human reproduction. In contrast with caffeine, PX has been approved by the Food and Drug Administration for administration to humans; it also has a higher hydrosolubility, which makes its use much easier than other methylxanthines (Tournay et al., 1995). It is considered to be a sperm movement enhancer (Tesarik et al., 1992a), hyperactivation agent, inhibitor of reactive oxygen species formation (Yovich, 1993) and acrosom reaction-improvement agent (Tesarik et al., 1992b). Ponce et al. (1999) showed that PX would preserve functional membrane integrity of mouse sperm tail. Addition of PX to the culture medium of mouse sperm caused completion of capacitation process (Duan and Goldberg, 2003). Others concluded that fertilization rate of mouse spermatozoa reduced after treatment with PX if sperm was not washed (Lacham- Kaplan, 1993; Trounson, 1993; Tournaye et al., 1994a,b). The aim of our study was to evaluate the ability of PX to enhance motility and fertilization rate of sperm samples after short time preservation in the different conditions. Materials and Methods Sperm preparation Spermatozoa were obtained from mature NMRI mice of proven fertility after separation from females for about two weeks. The animals were killed by cervical dislocation and spermatozoa were obtained from the cauda epididymides. The cauda epididymides were removed and immediately immersed into the T6 medium supplemented with 5mg/ml Bovine Serum Albumin (BSA)(Sigma, USA). Spermatozoa were released by mincing the epididymides in the collection medium. The sperm suspensions were incubated at 37°C in 5% CO2 for 1 hour. After incubation, sperm suspension was divided into 4 control and 4 experimental groups. Control groups included: (1) Fresh sperm sample (2) Preserved sperm sample at room temperature for 18 hours. (3) Preserved sperm sample at incubator 37°C for 18 hours. (4) Preserved sperm sample at 4°C for 18 hours. Experimental groups were treated by adding 3mmol/L PX to same as control groups. Assessment of sperm parameters The sperm parameters were assessed according to World Health Organization Criteria. The sperm parameters including: viability, motility, abnormal morphology, fertilization rate, cleavage rate and acrosome reaction. Viability: viability was assessed by eosin B (0.5 % in saline). A 20 µl sample of the sperm suspension was placed on a glass slide, mixed with 7 µl eosin, cover slip added and observed under a light microscope (× 400 magnification). Because of failure of plasma membrane, the head of spermatozoa absorb eosin and become red, but live spermatozoa remain colorless. Live spermatozoa were counted from each sample in five fields of vision randomly and percentage of live spermatozoa was recorded. Motility: A 20 µl sample of the sperm suspension was placed on a glass slide and observed under a light microscope (× 400 magnification). Motile spermatozoa were counted from each sample in five fields of vision randomly and percentage of motile spermatozoa was recorded. The grades of motility (from I to IV) were calculated, too. Abnormal morphology: Abnormal morphology was assessed using papanicolaou staining. With staining, the nuclei turn blue, acrosome and tail become pink. Abnormal morphology included loose head, bent neck and coiled tail. Abnormal morphology was counted from each sample in five fields of vision randomly and percentage of abnormal morphology was recorded. In-vitro fertilization (IVF): Oocytes were recovered from 6-8 week-old NMRI mice which had been super ovulated by means of intra peritoneal injections of 7.5 IU pregnant mare’s serum gonadotrophin (PMSG)(Nasr, Iran) followed 48 hours later by 7.5 IU human chorionic gonadotrophin (hCG) )(Organon, Holland). At 14 hours after hCG injection, the mice were killed by cervical dislocation and their oviducts were removed. Using a dissecting microscope, cumulus masses were released from the oviducts and placed into T6 medium supplemented with 15mg/ml BSA. Mature oocytes at metaphase ІІ were transferred into insemination dishes containing spermatozoa which had been treated according the experimental protocols added to insemination drops. The culture media were overlaid with paraffin oil and incubated at 37°C in 5% CO2. Fertilization rate was assessed 8 hours later by the presence of two pronucleus (2PN), and cleavage rate was assessed 24 hours later by the presence of two-cell stage embryos. Incubation with pentoxifylline A stock solution of PX (Sigma, USA) in distilled water was prepared each week and kept refrigerated at 4°C until used. Aliquots from this stock solution were added to the experimental tubes to achieve a final concentration of 3mmol/L. In preliminary dose-dependency experiments we showed that the maximal effect of PX on sperm motility characteristics had occurred when spermatozoa were treated with a dose of 3mmol/L (data not shown). Preparation samples for Transmission Electron Microscopy (TEM) Control and treated spermatozoa were fixed in 2.5% glutaraldehyde in phosphate buffered saline (PBS) for 2 hours at 4°C. The sperm samples were washed three times in PBS by centrifugationresuspension at 1000×g for 10 minutes and postfixed in 1% osmium tetraoxide for 1 hour. The samples were then centrifuged at 1000×g for 10 minutes and the pellets were dehydrated in graded acetone. Finally, the pellets were embedded in Epon. Thin sections were obtained with Reichert- Jug ultra microtome and stained with uranyl acetate and lead citrate, and observed with the transmission electron microscope to evaluate intact (with acrosome), reacting (with ruptured and irregular outer acrosomal membrane) and reacted sperm (without acrosome). Experimental Design The groups which were studied were as follows: Control group 1: Fresh mouse sperm Statistical analysis Survival after preservation, motility and abnormal morphology rates were analyzed using repeated measure and paired student’s test. The results of fertilization and cleavage rates were compared by x² analysis. The significant level was considered at P<0.05. Results The results are summarized in the tables 1, 2, 3 and 4, after sperm preservation at different temperature, the best survival rate was obtained at room temperature without significant difference with preservation at 4˚C (P > 0 .05) and significant difference with incubation at 37 ˚C (P = 0.002). The similar results were obtained after treatment with pentoxifylline. The motility rate was decreased significantly after preservation for 18 h and the best result was for maintenance at 4˚C and treatment with pentoxifylline (Table I). The progressive motility declined dramatically after preservation for 18 h and addition of pentoxifylline after storage at room temperature caused significant reduction (13.4 % without pentoxifylline versus 3.5% after treatment with pentoxifylline , P<0.05 ) (Table II). The most abnormal morphologies which were seen after preservation were bent neck, loose head and coiled tail. Treatment of preserved spermatozoa with pentoxifylline induced abnormal morphology significantly (P<0.05) (Table II). Table III shows fertilization and cleavage rates of control and experimental groups. Treatment mouse spermatozoa with pentoxifylline could improve fertilization and cleavage rates of fresh spermatozoa (45% versus 57% for fertilization rate, P<0.05 and 76% versus 815 for cleavage rate, P>0.05). Addition of pentoxifylline to preserved sperm at 4˚C improved the fertilization rate of 28% up to 52% (P<0.05) and the similar improvement of cleavage rate was obtained after incubation at 37˚C (58% before treatment versus 705 after treatment, P<0.05, Table III). Table IV shows the results of TEM study regarding acrosomal reaction. Treatment of sperm with pentoxifylline caused more spermatozoa to remain intact and less to react during preservation at the different temperatures. Discussion In the present study, treated spermatozoa with PX and preserved at 4˚C showed higher motility rate comparing to the other temperatures (room and 37˚C). Addition of PX to fresh spermatozoa increased progressive motility (grade IV) significantly. PX is a phosphodiesterase inhibitor similar to caffeine and can increase intracellular cyclic AMP levels (Numabe et al., 2001). Apraicio et al. (1979) demonstrated that the mean percentage of human sperm motility is apparently improved by in vitro incubation with PX. Further, it enhances a variety of human sperm- motion parameters (Tournay et al., 1995; Numabe et al., 2001). Ponce et al. (1999) observed an increase of motile sperm after addition of 5 mMol/L PX to mouse sperm. Gradil and Ball (2001) got the same results after addition of PXto equine spermatozoa. As Stanic et al. (2002) indicated cyclic AMP plays an important role in the glycolytic path way of the human sperm and, through its effect on glucolysis, it can influence the energy generation required for sperm motion. PX prevents cAMP breakdown by inhibiting the activity of the cAMPphosphodiesterase and, presumably, stimulates sperm motion. We added PX after preservation as Stanic et al. (2002) and Gradi and Ball (2001) did. Wang et al. (1993) concluded that post-thaw cAMP concentration in spermatozoa were found to be three times lower than pre-freeze concentration in the same samples, suggesting a low metabolic state in post-thaw spermatozoa. Duan and Goldberg (2003) showed that treatment of mouse spermatozoa with PX could complete capacitation. We did not evaluate the concentration of cAMP in mouse spermatozoa before and after preservation, but maybe addition of PX after short time preservation acts with a similar mechanism to postthaw addition which the others concluded (Gradi and Ball, 2000; Stanic et al., 2002). The results of the present study suggest that treating mouse sperm with 3 mM of PX is effective for mouse in vitro fertilization, especially after preservation at 4˚C for 18 h. Biological membranes have a liquid-like character and are often described as being fluid. A reduction in temperature causes the phospholipids bilayers to undergo a reversible transition to a more rigid gel (Melchlor and Steim, 1976). The capacitation-like membrane changes observed after cooling mouse spermatozoa were irreversible suggesting that they were not due to the lipid phase transition (Fuller and Whittingham, 1996). As some investigators note, (Fraser, 1987; Adeoya, 1993) capacitation involves a rise in the concentration of intracellular calcium. The change in membrane permeability during cooling may cause an increase in the concentration of intracellular calcium which initiates the capacitation-like changes seen in mouse spermatozoa after cooling (Fuller and Whittingham, 1996). The results also showed that preservation of mouse spermatozoa at 4˚C caused less cleavage rate and the reason could be the structural changes at cytoskeleton which has occurred during cooling. However we recommend further investigations regarding these issues. The purpose of TEM study was to evaluate acrosomal reaction precisely after preservation. However, the number of studied spermatozoa was not enough. The results showed that treatment of preserved spermatozoa with PX can prevent early acrosomal reaction (Table IV). Nasser et al. (1998) concluded that PX could enhance hyper activated motility at one hour and maintain this effect for up to 4 hours after its washing from the sperm suspension. Tesarik et al. (1992) also showed that the effect of PX persists for a few hours after its washing. The reason of improvement of fertilization rate of PX treated sperm and existence of more intact sperm at TEM study could be because of prolonged action of PX or as Ponce et al. (1999) indicated that in mouse sperm tail, PX would preserve functional membrane integrity, a relevant condition for adequate motility. PX may be considered an effective treatment to improve sperm penetration of oocytes after sperm maintenance at 4 ˚C for short period of time. However it is not clear whether PX induces capacitation and acrosome reaction of bull spermatozoa by an indirect or a direct mechanism. It is concluded that short time preservation can reduce the mouse sperm parameters and reduce its fertilization and cleavage rates. Treatment of mouse spermatozoa with PX can improve sperm parameters (survival and motility rates), fertilization and cleavage rates after short time preservation at 4ºC or 37ºC. PX also causes more sperm remain with intact acrosome. References
© Copyright 2004 - Iranian Journal of Reproductive Medicine The following images related to this document are available:Photo images[rm04011t4.jpg] [rm04011t2.jpg] [rm04011t1.jpg] [rm04011t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}