 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
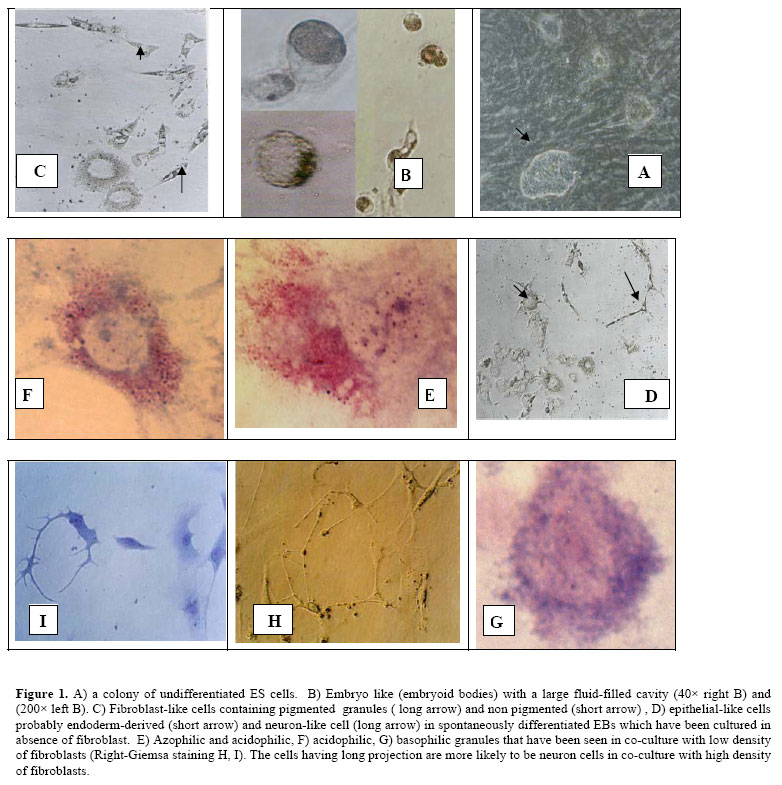
Iranian Journal of Reproductive Medicine, Vol. 5, No.1, Winter 2007, pp. 29-33 The effect of mouse embryonic fibroblast in direct differentiation of mouse embryonic stem cells Mahmoud Hashemi-Tabar, Ph.D., Fatemeh Javadnia, Ph.D., Mahmoud Orazizadeh, Ph.D., Ghasem Sakei, Ph.D., Maryam Baazm, M.Sc. Department of Anatomy, School of Medicine, Jondishapour University of Medical Sciences, Ahwaz, Iran. Correspondence Author: Dr. Mahmoud Hashemi-Tabar, Department of Anatomy, School of Medicine, Jondishapour University of Medical Sciences, Ahwaz, Iran. Received: 14 May 2006; Code Number: rm07006 Abstract Background: Since embryonic stem (ES) cells have the dual ability to proliferate indefinitely and differentiate into multiple tissue types, ES cells could potentially provide an unlimited cell supply for human transplantation. Key Words: Embryonic stem cells, Embryoid bodies, Mouse embryonic fibroblast, Coculture, Differentiation Introduction Embryonic stem (ES) cells are pluripotent cells derived from the inner cell mass of blastocyst-stage embryos that can replicate indefinitely in culture when maintained in an undifferentiated state by addition of maintained in an undifferentiated state by addition of leukemia inhibitory factor (LIF) to the culture medium (1,2). LIF implies suppression of differentiation and proliferation of ES cells through a cell surface complex composed of LIF receptor β (LIFRβ) and gp 130. These receptors can activate a transcription factor STAT3. Removal of undifferentiated ES cells from their feeder layer containing LIF and culturing them in suspension state, leads to the formation of embryo-like aggregates, known as embryoid bodies (EBs), in a similar process of cavitations that occurs in blastocyst stage embryos (3). Available data demonstrate that induction of ES cells into EBs and the cavitations of primitive endoderm in blastocyst are similar in signaling effects. The primitive ectoderm (epiblast) forms three primary germ layers: ectoderm, mesoderm, and definitive endoderm. These germ layers interact to form all tissues and organs of the developing embryo (4). The visceral endoderm is known to be an important source of patterning and differentiation signals in vivo (5). In addition, the ES cells in a similar signaling can differentiate into specific cell lineages that can be mediated by action of matrix components and/or growth factors (6). Molecular signatures including ligand/receptor and secreted inhibitors of different signaling pathways as fibroblast growth factor (FGF) and TGFb/bone morphogenetic protein (BMP) pathways are necessary to improve conditions not only to prevent differentiation but also to induce differentiation of ES cells (7). This will be extremely difficult since different growth factors have unique effects on the growth of undifferentiated ES cells and their differentiation (8). For example, one of the most routinely used mitogenic and growth factors for propagation of undifferentiated ES cells is basic FGF (bFGF) which has two distinct roles. Besides its mitogenic function on proliferating ES cells, and inhibition of endodermal differentiation in early phases, it can direct ES cells into ectodermal and mesodermal derivatives during the late courses of differentiation. It can also act as a differentiation factor for immortalised hypothalamic neuron stem cells (9). In vitro differentiation potential of mES cells and in particular, the use of human ES (hES) cells will facilitate studies on early human development and early embryogenesis events, which so far have been impossible (10). Their ability to spontaneously differentiate into a broad spectrum of derivatives of all three germ layers has also facilitated the examination of ES cells commitment to multiple cell lineages which will opens up a wide range of applications of ES cells technology, including preparation of an unlimited source of cells and tissues for transplantation therapy in various diseases and injuries (11, 12). However to date, the challenge remains to produce mature, functional and pure derivatives of ES cell that can be utilized for transplantation purposes. In this study, by comparing the cell morphology in the presence and in the absence of MEF cells, we have evaluated the differentiation potential of ES cells and studied the probable effects of factors that may be released from MEF feeder cells into medium. We then followed the progress underlying the directed ES cells differentiation in two systems. Materials and methods ES cells culture A previously established mouse ES cells in our laboratory was in 60 mm dishes containing inactivated MEF cells in 37oC, at 5% CO2 in a humidified incubator. ES culture medium was composed of Dulbecco's Modified Egals medium with high glucose (DMEM; Sigma Aldrich) supplemented with 10% FBS (Uroclone), 1% nonessential amino acids (Sigma Aldrich), 2 mM L–Glutamin (Sigma Aldrich), 0.1 mM 2-mercaptoethanol (Sigma Aldrich), penicillin/ streptomycin (100 U/ml and 100 µg/ml respectively, Sigma Aldrich); and 103 units/ml of LIF (Sigma Aldrich), which keeps ES cells in an undifferentiated state. Passage of ES cells on collagen coated dishes The undifferentiated ES cells was transferred to 24 well dishes (Cellstar) previously coated with 0.1% swine collagen type I (Sigma Aldrich) which had been treated with acetic acid 0.1% stock solution in 0.1 M, pH 7.2 PBS and cultured for one day in ES medium on the above condition to remove the MEF cells. Then the ES cells in each well were treated with mild 0.5% /0.5mM trypsin/EDTA (Sigma Aldrich) in PBS. The detached ES cells were diluted by ES medium without LIF in a density of 103 /ml and divided to three groups for EBs formation. Briefly the content of each well was transferred into the 4 well plates as follows. EBs formation in suspension and ES differentiation To induce the ES spontaneous differentiation in 24 wells dishes, ES cells were grown in the ES medium which contained 0.1% / 1mM trypsin/EDTA (sigma Aldrich) 20% FBS but no LIF. The ES cells in above condition were cultured in three different settings for 5 days to induce EBs formation: 1/ in the absence of MEF cells (group 1), 2/ in the presence of MEF cells with a density of 0.5 × 105 cells/ml (group 2), and 3/ in the presence of MEF cells with a density of 0.5 × 106 cells/ml (group 3). After 5 days, the supernatant containing EBs was collected from each well and were transferred into a new centrifuge tube for a few minutes to sediment the EBs. Sedimented EBs from each well were then transferred to the collagen-coated 4-well dishes to attach to substrate and were cultured for 20 days. The medium was changed every 4-5 days. Spontaneously differentiation of ES cells was checked every day by an inverted microscope. In order to detect differentiating haematopoeitic cells, ES cells and their derivatives were examined by morphology. They were also fixed (by acetic acid and metanol 1:3) and stained by Wright-Gimsa method. Results In a standard ES cell culture system, there may be numerous different cytokines and growth factors which can be released from MEF feeder layer. In this study, the possible effect of MEF cells on ES cell differentiation was investigated. In general, the undifferentiated mES cells with distinct colony boundaries but with indistinguishable cell boundaries (fig 1-A) have a potential to differentiate into EBs if they are cultured in suspension state. During 5 days of suspension culture, mES cells loosely aggregate and begin to form a large fluid-filled cavity. Embryo-like EBs appeared in all of three studied groups (fig 1 B). Appropriate number of EBs from each group was plated on collagen-coated dishes and was checked every day for attachment to the substrate by an inverted microscope. In group 1, various cellular structures displaying epithelial-like, fibroblast-like and nerve-like morphologies were seen in spontaneously differentiated EBs (fig 1 C, D). In group 2 in which mES cells were cultured with low density MEF cells, numerous population of cells containing azophillic, acidophilic and basophilic granules was observed ( fig 1 E,F,G). In cells that contained acidophilic granules, nuclei were either intact or degenerated (fig 1 E, F). An heterogeneous population of cells including neuron-like cells with long projection-like extremities ( fig 1H,I) and fibroblast-like( fig 1I) cells were observed in group 3, in which mES cells were cocultured with high density MEF cells. Epithelial-like cells morphology was not observed in group 2 and group 3. Discussion Pluripotent ES cells are capable of differentiating into many cell types. The promising of using these cells for medical applications, including cellular transplantation, has attracted more attention in the field of ES cells isolation and differentiation (13). In 1985, the blood formation from mouse ES cells was reported in vitro (14), but achievement of stable blood engraftment in irradiated mice with ES-derived progenitor cells still remains a challenge. Limited success in hematopoietic engraft deserves more investigation in the field of transcription factors and other soluble molecules to delineate the underlying mechanisms (14). The main goal of this study was to examine the conditions which are required by mouse ES cells to produce a uniform population of precursor cells or fully differentiated cells in vitro. Previous work with human ES cells showed that several types of cells can be enriched in culture either by the addition of growth factors or by introduction of transcription factors (15-17). Whereas conditions that direct differentiation of mouse ES cells into different phenotypes such as; neuronal, hematopoietic, and endocrine have been published (18). In this study, we have demonstrated that lack of MEF releasing factors promote spontaneous differentiation of ES cells derivatives of all three germinal layers including; epithelial like cells that arise from endoderm (i.e. hepatic cells and pancreatic cells derived from ES cells), fibroblast like cells derived from mesoderm (i.e. smooth and striated muscle cells haematopoietic lineages), and neuron-like progenitor cells. In the present of low density MEF cells, the ES cells directed to hematopoeitic cells with different azo, acidic and basophilic granules based on the Romanowsky staining method. It is useful to use PCR and RT-PCR for detection of gene expression pattern such as GATA-1,2 and FOG-1 that is essential for hematopoietic differentiation and EBs differentiation to hematopoietic precursors (19). A possibility to identify growth factors is to analyse feeder-cellconditioned medium. Lim and Bodnar (2002) identified 136 unique protein species such as IGF-binding protein 4, pigment epithelium-derived factor, SPARC (secreted protein, acidic and rich in cysteine), nidogen-2, galectin 1 and peroxiredoxins by analyses of conditioned medium from MEF cells, and the STO cell line (20). The growth factors based on their differentiation effects can be divided into three groups: The first group (TGF-b1 and activin-A) appears to inhibit endodermal and ectodermal cells, but allow differentiation into mesodermal (muscle) cells. The second group includes factors that allow or induce differentiation into ectodermal as well as mesodermal cells (RA, bFGF, BMP-4, and EGF), whereas the third group (NGF and HGF) allows differentiation into all three embryonic lineages including endoderm (9). Differentiation of ES cells in a serum-free system demonstrates that BMP-4, but not several other related TGFb family members, induces hematopoietic cells from primitive mesoderm in EBs. BMP-4 induces ventral-type mesoderm formation, including blood cells, in Xenopus embryos (21). Therefore, our results demonstrated that differentiation of ES cells is a dose-response process, depending on factors that may be released into the ES cell medium. BMP-4 indeed is one of the most important factors which involve in haematopoeiteic differentiation from mesoderm. It is very likely that the concentration of BMP-4 that is required for ES cell differentiation into haematopoeitic cells is optimal in low density of MEF cells. It is unknown whether this factor acts on pre-hematopoietic mesoderm or on committed hematopoietic cells. Thus, ES cells in this experiment can be used to test candidate growth factors for initiation and support of embryonic hematopoiesis. EBs themselves also offer a potential source for new cytokines that stimulate primitive and/or definitive hematopoiesis. Therefore in the presence of low density MEF cells, mES cells differentiate towards hematopoietic lineage and produce relatively large numbers of early embryonic hematopoietic precursor cells. These precursors can be isolated and manipulated for further studies in the field of embryonic blood cell development. Conclusion Differentiation of mES cells into derivatives of three germ layers is a dose response process, depending on factors that are released from MEF feeder cells into ES media in a coculture system. Our experimental approach shows that the establishment of a system in which mES cells are induced to produce embryonic hematopoeitic cell lineages for investigational proposes can be very valuable for further research on early hematopoiesis and the analysis of hematopoeitic precursors. If both primitive and definitive hematopoiesis cells are aroused from one precursor cells in an EB, it could be very feasible to isolate and characterize this precursor from early EBs for future studies which investigate the mechanisms responsible for the derivation of haematopoietic precursor’s cells. References
© Copyright 2007 - Iranian Journal of Reproductive Medicine The following images related to this document are available:Photo images[rm07006f1.jpg] |
| |||||||||

{kind=link}