 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Iranian Journal of Reproductive Medicine, Vol. 5, No.2, Spring 2007, pp. 45-50 The effect of hyaluronic acid on motility, vitality and fertilization capability of mouse sperms after cryopreservation Mitra Bakhtiari1 M.Sc ., Aligholi Sobhani1 Ph.D., Mohammad Akbari1 Ph.D. , Parichehr Pasbakhsh¹ Ph.D., Mehdi Abbasi¹ Ph.D., Azim Hedayatpoor¹ M.Sc., Fardin Amidi² Ph.D., Feridoon Sargolzaei3 Ph.D. 1 Department of Anatomy and Vali-e-Asr Reproductive Health Research Center, Faculty of Medicine, Medical Sciences/University of Tehran, Tehran, Iran. Correspondence Author: Dr Aligholi Sobhani, Department of Anatomy and Vali-e-Asr Reproductive Health Research Center, Faculty of Medicine, Medical Sciences/University of Tehran, Tehran, Iran.E-mail: sobhania@tums.ac.ir Received: 27 September 2006; accepted:5 March 2007 Code Number: rm07011 Abstract Background: Various approaches have been used in the attempts to
improve the quality of frozen–thawed mouse sperms. According to
literatures, it seems that hyaluronic acid (HA) has an important role on the
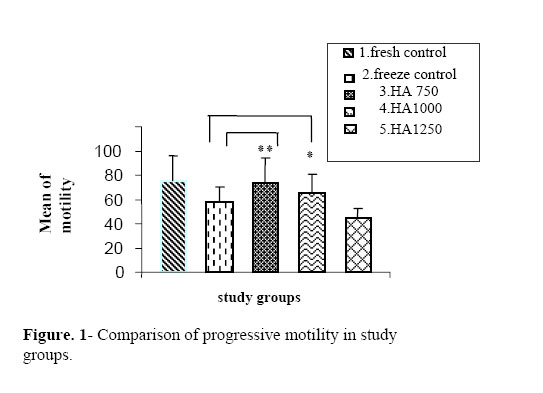
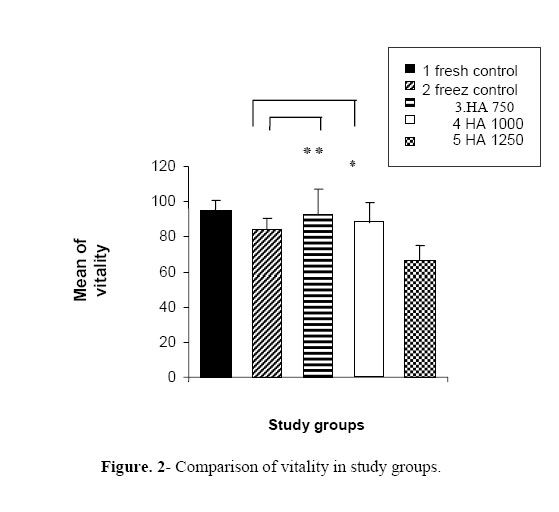
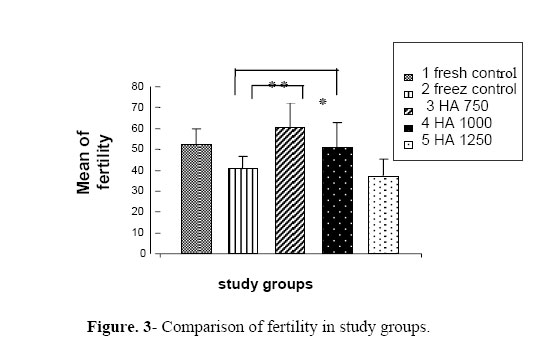
permeability and motility of sperms and their interaction with gametes. Key words: Hyaluronic acid (HA), Cryopreservation, In vitro fertilization (IVF), Sperm parameters. Introduction Recently, a large number of in vitromutations have been induced in various mice strains, world wide (1-5). As a result, the number of mice strains with induced mutations is rapidly expanding, and the maintenance of these strains by standard colony breeding is becoming an increasing problem. Embryo freezing is generally used for this purpose (6). However, in the conventional method of embryo freezing, 500 embryos per strain are required from the oviducts of 20–50 mated females or by in vitro fertilization before the freezing procedure (7). In contrast, 10–30×106 spermatozoa can be frozen immediately after collection from the epididymes of each male. If all frozen-thawed spermatozoa from one male are used for in vitro fertilization, they can fertilize at least 500 oocytes (8). Therefore, sperm freezing may provide a much simpler and economical alternative to embryo freezing (9). Freezing and thawing induces a number of insults to the sperm cell, such as cold shock, osmotic stress and intracellular ice crystal formation, which may kill or grossly damage the spermatozoa (10). Various approaches have been used in the attempts to improve the quality of frozen-thawed mouse sperms, including the changes in holding-time before freezing (11), and the addition of various additives (12, 13). Glycosaminoglycans (GAGs) are essential components of the extracellular matrix, contributing to the cell recognition, cellular adhesion and growth regulation. The GAGs can be classified by their sugar residues and other characteristics such as, sulfation into four main groups of GAGs (14) including the non-sulfated glycosaminoglycan and hyaluronan (HA). Important aspects of sperm functions such as, motility (15, 16) and capacitation (17, 18) appear to be mediated, at least partially, through HA. Addition of HA to fresh (19) and frozen–thawed sperms (20) improves sperm motility. Interestingly, HA not only improves sperm motility after freezing and thawing procedures, but it further appears to stabilize the spermatozoa with already declining motility in frozen/thawed sperms (20). Exogenous HA decreased polyspermia during conventional porcine IVF (17), an effect probably related to the capacity of HA to delay sperm capacitation and/or the acrosome reaction (17, 18). The aim of the present study was to test the hypothesis that supplementing with HA has an important role on the viability and motility of sperms and the interaction of gametes with these sperms. Therefore, we assessed the effect of different doses of hyaluronan on the motility, vitality and fertility rate of mice after cryopreservation. Materials and methods Animals In total,6 Adult NMRI mice (8–10-weeks old) were obtained and maintained at 20 ºC in a fixed 12 hours light and 12 hours dark cycles with free access to the food and water. Cryoprotectant agent (CPA) At once, 9 ml H2O was placed in a screw top 15 ml Falcon tube and equilibrated to 60 °C in a water bath. Then, 1.8 g raffinose (Merck, Darmstadt, Germany) followed by the addition of 0.3 g skimmed milk (USA Difco Betalab) and dissolving by gentle inversion. Then, the mixture was aliquoted into microfuge tubes and centrifuged at 14,000 rpm for 10 min. Then the supernatant was collected and filtered by using a 0.45 µm syringe end filter and Placed in 1.1 ml aliquots into cryo-tubes and stored at –20 °C. Cryopreservation method To prepare the cooling apparatus, a platform was placed (e.g. the insert from a Gilson yellow tip box) into the polystyrene box. This acts as a support for the cryotube rack. The liquid nitrogen was carefully poured into the polystyrene box to just cover the platform. A cryotube rack was placed on the top of the platform so as to make it suspended in liquid nitrogen vapors. The lid was replaced on the polystyrene box for permitting it to fill with vapors. The liquid nitrogen was replenished as necessary during the freezing session, but the level was not allowed to rise above the platform.One 1.1 ml aliquot of cryoprotectant solution for each male mouse was thawed and brought to 37 °C in the incubator to disperse the sperms; the culture dish was gently shaken for ~30 seconds. The adult mouse was killed by cervical dislocation. Cauda epididymes was removed following the aseptic cleaning of all the fat and blood. The cleaned organs were transferred to the dish and, by using a pair of watchmaker’s forceps and micro-spring scissors, the epididymes was minced and spermatozoa were dispersed into the CPS by shaking the dish for about 2 min and incubated for sperm preparation for 10 minutes at 37 °C.Then, the animal tissues were removed from the suspension by scraping them to one side of the culture dish with a pipette tip. Aliquotes of 100µl were placed into each of 8 cryotubes and the screw cap was tightened to seal them. The cryotubes were placed in a pre-cooled rack, supported in the liquid nitrogen vapor phase of the freezing apparatus and was left for 10 minutes. The cryotubes were removed from the freezing apparatus and plunged into the liquid nitrogen. The tubes were then stored in a liquid nitrogen refrigerator until required. Thawing of spermatozoa By using forceps, the cryotubes were held in the air for 30 seconds and then thawed rapidly by plunging them into a 37 °C water bath. When the sample was thawed, the cryotube were gently agitated to ensure that the sperm sample is evenly mixed. Subsequently, the sample was centrifuged at 5000×gfor 5 min. The supernatant (cryoprotectant) was discarded and replaced with human tubal fluid (HTF). The sperm sample was gently re-suspended by tapping the tube with fingertips and incubated for 10 min at 37 °C to allow a minimal selection by “swim up”. Sperm HA-supplementing experiments The sperms of each mouse were divided into five groups. Group 1 (fresh control): These samples were not frozen and maintained in HTF media without any hyaluronic acid supplementation for 2 hours. Group 2 (frozen control): they were maintained in HTF media without any HA supplementation for 2 hours after thawing. In groups 3, 4 and 5, HA with the doses of 750, 1000 and 1250µg/ml were added into HTF media after thawing, respectively. After 2 hours of incubation, the sperms were assessed. Sperm analysis Motility parameters were measured by using binocular microscope. The sperms were divided into 4 groups of motility (according to WHO category); grade 1: rapid progressive, grade 2: slow progressive, grade 3: non-progressive and grade 4: immotile. Toanalyze viability, we used a vital staining, eosin B and determined 100 cells per sample. The viable cells had green head and dead sperms appeared red in color. IVF methods Female mice (8-10 week old) were super-ovulated by 10 units of PMSG followed by HCG administration (10 units) after 48 hours (intraperitoneal injection). After 14 hours, the female adult mice were scarificed by cervical dislocation, then fallopian tube was removed aseptically and tissue was punctured under dissection microscope. Oocytes were removed by mouth micropipettes in metaphase II stage into IVF medium. In all groups, 10 oocytes in metaphase II stage were put in each IVF drop. Then swam-up sperms were removed after capacitation (1.5 hours in Ham’s F10 + 4 mg BSA) and were added to oocytes in IVF drops (T6 + 15 mg BSA) followed by incubation. Oocytes were washed after 6 hours and removed to IVC drops (T6 + 4 mg BSA). Then, two-cell stage embryos were assessed after 24 hours. Statistical analysis To compare the various groups was used student’s t-test. The entire statistical analysis was carried out using the Statistical Package for Social Sciences (SPSS for Windows). Statistical significance was set at p < 0.05. Results In this study, we considered the progressive motility only. In group 1 (fresh control), the motility was 75.8%, in group 2 (freeze control), it was 58.06%, and in supplemented groups 3, 4 and 5, the progressive motility was 73.80%, 64.45% and 44.10%, respectively. Therefore, 750 and 1000 µ g/ml HA had positive effect on progressive motility. The effect of 1250µ g/ml HA was negative on the improvement of motility (Fig. 1). In group 1 (fresh control), and 2 (freeze control), the vitality rates were 95.03% and 84.30%, while in supplemented groups 3, 4 and 5, the vitality was 93.05%, 87.70% and 67.05%, respectively. Therefore, 750 and 1000 µ g/ml HA had positive effect on the percentage of viable sperms. The effect of 1250 µ g/ml HA was negative onsperm vitality improvement (Fig. 2). In group 1 (fresh control)and 2 (freeze control), the fertility rates were 52.06% and 40.65%, respectively while, in the supplemented groups 3, 4 and 5, the fertility rates were 60.20%, 51.10% and 37.30%, respectively. Therefore, 750 and 1000µ g/ml HA had positive effect on fertility rate of sperms. The effect of 1250µ g/ml HA was negative on the improvement of fertility rate (Fig. 3). Discussion Rodent sperms have proven to be more difficult to cryopreserve than other mammalian sperms. Rat sperm cryopreservation has not yet been accomplished and mouse sperm cryopreservation was reported only 10 years ago. Difficulties in reproducing the original results (21, 22) inspired modifications (23, 24) to protocols that make freezing morereliable but still are not equally successful for all mouse strains.Classic cryobiology studies suggest the use of supplement to efficiently protect the cells from freezing injuries through colligative or “solution effects” (25). This principle has been empirically applied in mammalian sperm preservation since early 1950 (26). It seems that the choice of supplements is a matter of trial and error, because the action of these is still not totally understood (26). Poor fertility was reported for mouse sperms frozen in 18% raffinose and 3% skimmed milk despite the good recovery of sperm motility (27). Hyaluronan plays a major role in important physiological functions of the spermatozoa, such as capacitation (28) and motility (16). Recently, Huszar et al (2006) reported that Hyaluronic acid mediated sperm selection is a novel technique that is comparable to sperm-zona pellucida binding (29). In the present study, HA-supplementation, after thawing, yielded higher sperm motility values post-thaw. This observation is similar to that previously described for frozen/thawed human semen, where HA-addition after thawing resulted in the significant increases in sperm motility and velocity (20). In the experiment of Sbracia et al (1997), HA was added to a final concentration of only 50 μg/ml (20). It may then be possible that the higher doses used in our experiment allowed the maintenance of an effective and unaltered concentration of HA after thawing. Alternatively, in addition to possible innate species differences, the difference may be due to differences in the levels of supplementation. We added HA to a final concentration of 750, 1000 and 1250 μg/ml, while HA-supplementation yielded a significant improvement of post-thaw motility. The result of present study emphasized that the hyaluronan may have an influence on motility, vitality and fertility rate of sperms and the dose of 750µ g/ml had a significant effect on these factors. The mechanism behind an HA-induced increase in sperm motility remains to be determined. An HA-receptor has been localized along the tail, mid piece and head of human spermatozoa (30). Blocking this receptor with mono-specific antibodies resulted in the inhibition of sperm motility. Moreover, enhanced phosphorylation of the 34 kDa HA-binding protein (HABP) has been observed in response to HA supplementation, suggesting an important role of this glycosaminoglycan in initiating the signal transduction controlling the important processes of sperm physiology, such as motility (31, 32) that may result in an increase of the ATP levels and improved flagellar function (20). The human study found no effect of HA when supplemented prior to cryopreservation and it was suggested that the freezing/thawing procedures cause deterioration of the HA structure. The pre-ovulatory sperm reservoir in the oviduct contains large amounts of GAGs, including HA (34) that possibly maintain sperm viability during their residence in the reservoir (34). It is possible that HA is inhibiting capacitation, similar to the in vitro effect observed in the present study (35). Since HA improves post-thaw sperm motility and viability, and given its presence during sperm–oocyte interactions in vivo (28, 36), we suggest that HA-supplementation is a safe and readily applicable approach to improve the quality of frozen–thawed mouse sperm. In another study, the supplementing extended boar semen with 500 or 1000 μg/ml HA prior to the freezing on post-thaw sperm motility parameters, and membrane lipid architecture status, showed that HA appeared to preserve post-thaw spermatozoa viability in vitro and maintained membrane stability after cryopreservation (21). Concurrently with the sperm maturation studies, in another line of experiments, the effect of HA was investigated, which is a linear repeating polymer of disaccharides, on human sperm function. Hyaluronic acid in the medium increased the velocity and retention of motility and viability in freshly ejaculated, as well as in cryopreserved–thawed human spermatozoa (37). The enhancement of sperm motility and velocity occurred as a direct response to HA, as indicated by two observations: 1- there was an instantaneous increase in sperm velocity and tail cross-beat frequency upon HA exposure and 2- when, after density gradient centrifugation, HA-exposed sperms were tramsferred to the standard medium, the motility and velocity properties returned to those of the control sperms. They concluded that the effects of HA on sperms are likely to be receptor-mediated, in line with the evidence, established by various laboratories for the presence of the HA receptor in human sperms (38- 40). In this study, 750 μg/ml HA had a significant effect on motility, viability and fetility rates. The dose of 1000 μg/ml HA had positive effect on the improvement of sperm characteristics but 750 μg/ml was the best choice. In contrast, 1250 μg/ml HA reduced motility, viability and fertility rates. This matter can be described with HA-binding receptors on sperm membrane. In our investigation, none of these doses had an effect on sperm morphology. Anyway, Huszar et al (2006) suggested a relationship between HspA2 expression in elongating spermatids and events of late spermiogenesis, such as cytoplasmic extrusion and plasma membrane remodeling that aid the formation of the zona pellucida binding and hyaluronic acid binding sites (29). Conclusion We suggest that using 750 μg/ml HA after thawing the mouse cryopreserved sperm can obtain better IVF results. However, it is suggested to investigate the effects of HA in other species, especially in human. Acknowledgements We thank research deputy of Medical Sciences/University of Tehran for the financial support during this work. References
© Copyright 2007 - Iranian Journal of Reproductive Medicine The following images related to this document are available:Photo images[rm07011f3.jpg] [rm07011f1.jpg] [rm07011f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}