 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Iranian Journal of Reproductive Medicine Vol. 5, No. 4, Autumn, 2007, pp. 151-157 Non-invasive evaluation of embryo morphological plasticity by designing of new transgenic gene cassette Abdolhossein Rezaeian1,2M.Sc., Seyed Mehdi Kalantar1Ph.D., Safar Farajnia2M.Sc., Mehrdad Soleimani 1 B.Sc., Abbas Baghi1 B.Sc., Abbas Aflatoonian1 M.D., Sirous Zeinali2 Ph.D. 1 Research and
Clinical Center for Infertility, Shahid Sadoughi University of Medical Sciences,
Yazd, Iran. Received: 15 September 2006; accepted: 5 August 2007 Code Number: rm07030 Abstract Background:
Determination of transgenic embryos from non
transgenic embryos sibling is an important step in producing homozygous
transgenic mice. These steps need by PCR or southern blotting
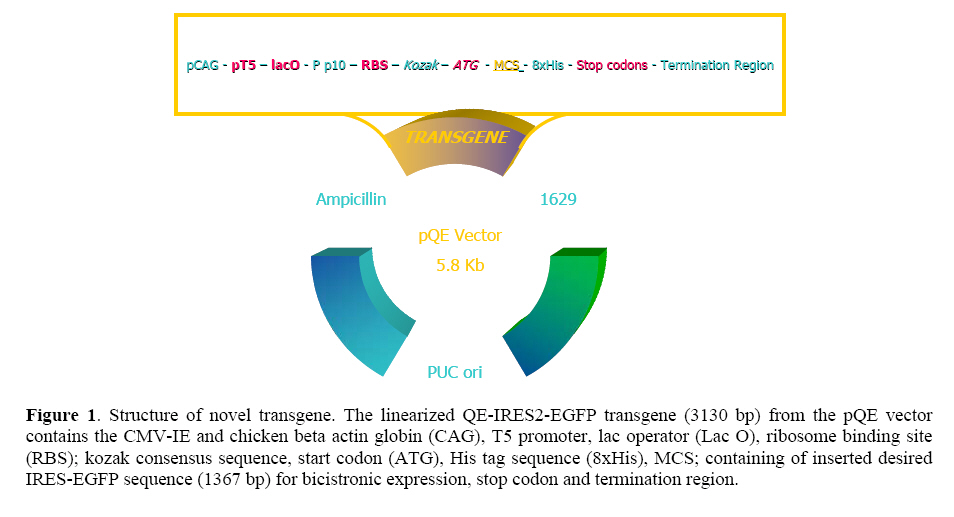
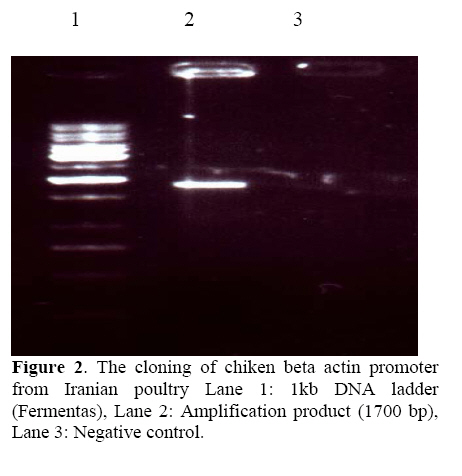
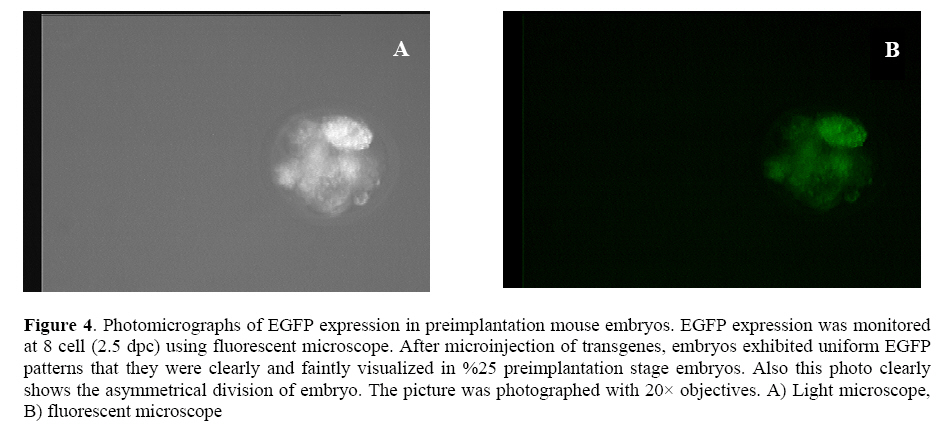
followed extraction of DNA, but both techniques require skill and consume time. Key words:Enhancedgreen fluorescent protein (EGFP), Transgenic animals, Embryo. Introduction Since the first report of the green fluorescent protein (GFP)expression in Caenorhabditiselegans (1), this novel and simple reporter has attracted much interest as an in vivo marker for gene expression. Other gene reporters, which encode the specific enzymes firstly require substrates to detect the gene expression, but the detection of GFP does not require the extraction, substrate loading steps and following toxicity, so we can monitor the presence of GFP by illuminatin living cells. Already, several substrates have been developed that can be permeable to the plasma membrane and are processed in the cytoplasm for use in living cells (2). However, it is still necessary to load the substrate with thesemarkers (3-5). On the other hand, the discernment of transgenic embryos from non transgenic embryos sibling is an important step in producing homozygous transgenic mice. These steps need by PCR or southern blotting followed extraction of DNA (4, 6) but both techniques require skill and consume time. Thus, to circumvent these problems, it seems that selection of embryos before implantation with an integrated transgene for embryo transfer will have high efficacy in transgenic technology regardingthe long gestation period, limited number of offspring and low percentage of implantation if applied to a large number of transgenic animals. After the production of the founder transgenic animals, it would be necessary to propagate the transgenic animals. Therefore, for the production of large transgenic animals such as cow andpig, the detection of the transgenes at the preimplantation stage would be desirable. Therefore, the method for quick separation of transgenic embryos with high quality will be important for the rate of implantation after embryo transfer.In addition, blastocyst biopsy and subsequent PCR analysis enable us to separate X and Y chromosome bearing embryos (1, 5) Which will be important in animal breeding and the production of milk and meat. Several reports have suggested the possibility of using marker genes for this purpose (7, 8). In this context, we describe the designing of novel transgene, which enables us to separate quickly and non-invasively high quality transgenic embryos and also parallel monitoring of the effect of exegonous geneon mammalian cells. We demonstrate this applicability by designing the novel GFP transgene. Moreover, we try to assay morphological plasticity of the produced transgenic embryos after microinjection system to evaluate this new transgenic gene cassette and its usefulness probably in livestock industry. Materials and methods Animals The outbred six-week old CXB RI females were resulted by C57BL/6 ~ BALB/c mating from the breeding farm of Pasture Institute of Iran. Pregnant female mice were sacrificed humanely for embryo biopsy by cervical dislocation method (9). Construction of EGFP expression vectors In brief, the CAG promoter, which was composed of the CMV enhancer, a fragment of thechicken β-actin promoter and rabbit β-globin intron was used. The IRES-EGFP fragment covering the coding sequence from plasmid pIRES2-EGFP (Clontech, Palo Alto, CA) was digested with BglII and NotI and inserted into the pQE-Tri system expression vector (Qiagen, Hilden, Germany) containing the CAG promoter and rabbit β globin termination region. Transgenes were excised immediately downstream of the polyadenylation signal and upstream of the CMV-chickenβactin promoter with the PaeI and Esp3I enzymes and the entire insert was gel purified (QE-IRES2-EGFP; 3.1 kb). For construction of another GFP expression vector used as control, in brief the chicken β actin promoter was amplified by PCR with 5' primer; TACGTATCGAGGTGAGCCCCACGTTCT and 3' Œ primer; GCTAGCCGGTCACACGCCAGAAG from the genomic DNA of Iranian poultry (Research Center of Animal Science, Karaj, Iran) as a template. SnaBI and NheI sites included in the PCR primers were used to introduce this amplification product into pIRES2-EGFP instead of the short chicken β actin promoter sequence that there was in the first designed construct. The entire insert with the promoter and coding sequence was excised with AseI and AflII and gel-purified (Qiagen,CA)(Figure 1). Microinjection and production of transgenic embryos The outbred six-week old CXB RI females were superovulated by injecting 5IU of PMSG and hCG, respectively, at 48 h intervals and then fertilized one-cell embryos of CXB RI females mated with CXB RI males were collected from the oviduct. Transgenicgreen embryos were produced by microinjecting the purified PaeI and Esp3I fragment with QIAEX (QIAGEN, CA) into male pronuclei of recovered fertilized eggs. This procedure was also performed using AseI and AflII fragmentresulted from the second construct. After microinjection (Nikon 300, Hoffman) of EGFP transgenes, embryos were cultured. Preparation of embryos and in vitro fertilization (IVF) On the day following hCG injection and for sperm mediated gene transfer using IVF technique, mature oocytes (MII) were recovered by opening the oviducts in pre-warmed M16 medium supplemented with 10% bovine serum albumin. At the same time, mature male CXB RI was killed to remove the tail of epididymis. The extracted spermatozoa were then cultured in M16 droplets for 2h at 37ºC, 5%CO2. The IVF took place approximately 5 h post sperm incubation. Observation and selection of embryos For quality prediction of preimplantation embryos after injection using our novel transgene, three major standard criteria were used; 1- the rate of development, 2- the degree of fragmentation, 3- the intensity of EGFP expression (10). Also to evaluate the ability of development to blastocyst (4 dpc) and viability of transgenic embryos, EGFP-transgene injected (EGFP treatment), T10E0.1 buffer injected (physical treatment) and non-injected IVF eggs (medium treatment) were cultured. The incorporation and separation of green and non green embryos was examined by placing step by step developing embryos under UV light usinga Zeiss (Germany/Axiovert) inverted fluorescence microscope withfilters BP 450-490, FT 510, LP 520. EGFP was distinguishable by shedding the different excitation light. The eggs expressing EGFP emit green when excited with 488 nm and detected at 500-to 530 nm band pass filter. Separated embryos were washed and cultured in M16 medium for development and further assay using PCR analysis. PCR amplification of embryo samples One blastocyst was transferred into an Eppendorf PCR tube with 10 μl of dH2O, overlaid with 30μl of light mineral oil. The samples were heated for 10 min at 97◦C. After cooling down, 1μl of proteinase K stock solution (10 mg/ml) was added to each tube, and then treated for 1 h at 56◦C. Proteinase inactivation was done for 10 min at 97◦C. Then 3.5 μlof dH2O, 2 μl of 10X PCR buffer (100mM Tris-HCl, pH 9.0, 500 mM KCl, 1℅ Triton X-100), 1.5 μl of MgCl2 stock solution (25 mM), 1μl of dNTP mixture (each 5 mM of dATP, dCTP, dGTP, dTTP) and 0.5μl of each primer (50 μM) were added to individual tubes. The primers used for the detection of the GFP were: 5Œ TGGAGAGGGTGAAGGTGATGC and 3' TGTGTGGACAGGTAATGGTTG. Finally, 5μl of polymerase solution (2.5 units of Taq polymerase in 1~PCR conditions) were added. Therefore PCR reaction was performed by:40 cycles of denaturation at 94ºC for 30s, annealing at 54 ºC for 30 s and extension at 72 ºC for 40 s. After the completion of 40 cycles, 5 μl of each sample was analyzed immediately by 1% agarose gel electrophoresis. Protein analysis The DNA construct was transformed into E. coli strain JM109. An overnight culture of bacteria (5 ml) was diluted into 300 ml of LB medium. At OD 600 = 0.4, the culture was induced by IPTG (1 mM) and incubation was continued for 4 hr at 37◦C. Cells were then pelleted and resuspended in 10 ml of Sonication buffer (SB 50 mM Na-phosphate PH 7.8, 300 mM NaCl) and after washing with PBS were lysed by sonication (6 times for 30 s in an ice bath). The lysate was clarified by centrifugation at 14000 rpm for 20 min and applied to 1 ml Ni-NTA (Qiagene) column equilibrated with SB (pH 8). The column was washed with 50 ml of SB (PH 6) and the protein eluted with 8 ml of SB (pH 4.5). SDS-PAGE method was performed under non-reducing conditiones using a 12% gel. Gels were stained with 0.1%Coomassie brilliant blue R250 (Biorad, Richmond, CA) and destained in isopropanol:acetic acid:water (1:1:8) by gently shaking at RT. Results Expression of EGFP in preimplantation mouse embryos Of 72 embryos microinjected with novel EGFP transgene, 69 survived and were cultured for further development. After 4 days in vitro culture fallowed by microinjection, 44 embryos developed to blastocysts. In this period, EGFP was easily detected by fluorescence microscopy at two-cell, to morula and blastocyst stages in 17 embryos (EGFP-positive) while embryos 27 were EGFP-negative. The percentage of survived embryos to developed blastocyst in EGFP expressing embryos was less than physical and medium treatment (IVF) groups. Also in our transgenic embryos the percentage of EGFPpositive embryos from surviving embryos after microinjection was remarkablely higher than most published data which means creating of good morphological plasticity (Table I, Figure 2). Table I. Microinjection experiment with novel transgene and CXB FI superovulated eggs. a) Positive samples; EGFP gene treatment embryos, b) T10E0.1 buffer injected embryos; physical treatment and c) non-injected embryos are for evaluation of medium effect.
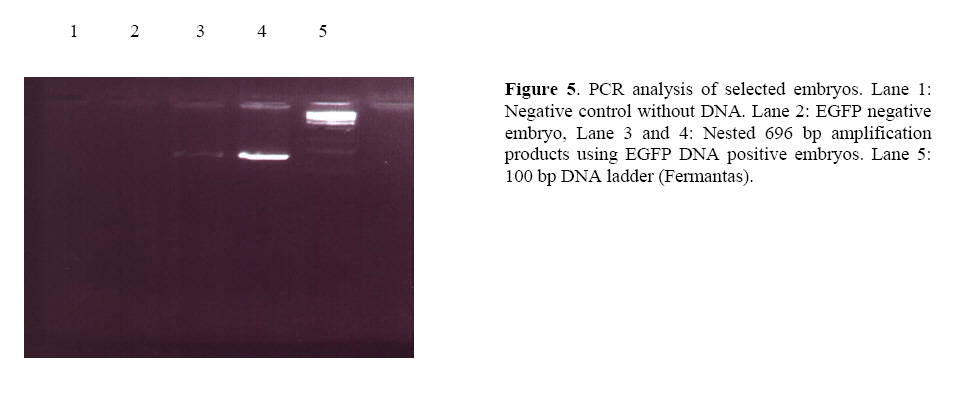
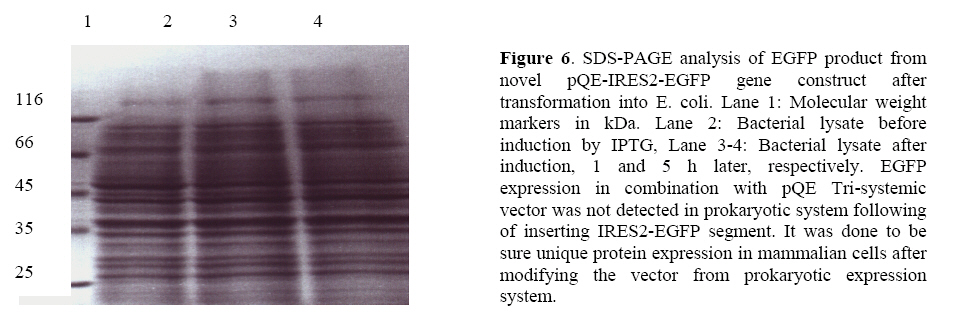
Transgenesis identification from EGFP expressing blastocyst To verify the amplified sequence and increase sensitivity, we performed two rounds of PCR analysis firstly using EGFP-positive blastocyst genomic DNA and secondly EGFP transgene-specific primers. On the nested PCR analysis, transgene specific 696 bp bands were amplified from first round PCR amplification proved that all the selected green embryos inherited EGFP transgene. No amplification was detected with genomic DNA from EGFP-negative embryos (Figure 3, 4, 5). Protein analysis After sub-cloning of IRES-EGFP sequence into pQE Tri systemic vector, we didnft see the evidence of EGFP protein expression into bacterial cells in which it was detected before ligation of IRES segment with EGFP gene. The SDS-PAGE experiment confirmed this phenomenon. This was additional confirmation to check the accuracy of EGFP expression pattern following of sub-cloning and changing of expression shift into mammalian cells. The reason was the existence of IRES sequence, which had made a long distance disruption between RBS and start codon of EGFP gene in bacterial cells (Figure 6). Discussion The aim of this study was simulation of high accuracy method using novel EGFP gene cassette to eliminate some consume time in livestock industry and high quality embryo morphological selection. Southern blotting procedure requires skill and it consumes time if applied to detect a transgene between large numbers of transgenic animals. Also the low efficiency of transgenic animal production by microinjection has been a serious problem, especially in large mammals because of long gestation period and limited number of offsprings. Already the enhanced green fluorescent protein (EGFP) has been known the best marker whose codon usage has optimized for human cells (11). The preliminary usage of substrate is not imperative in EGFP gene reporter. Otherwiseusing of EGFPmutant that has an excitation peak at 490 nm, minimized damage to embryos by ultraviolet light (12). Thus non-invasive selection of transgenic embryos was performed successfully at the pre-implantation stage of embryos using EGFP gene marker under our novel pQE-IRES-EGFP-Tri systemic gene cassette (Figure 4). We modified pQE-Tri systemic vector (Qiagen) with EGFP and IRES sequence to trace out comingmolecular-farming gene by co-injection method. It means by available GFP variants, which are different in spectra, we are able to study the characters of another co-injected and expressed gene in developmental research area. This novel transgene with restricted role to play bicistronic expression for both desired genes using IRES, also will be suitable for pharmaceutical planningwhen be co-injected with second gene. It is able to assay every gene after a little modification using three specific promoters located in pQE vector, into three separated prokaryotic, baculovirus and eukaryotic systems. Furthermore, we replaced the chicken beta actin promoter colonized from our poultry in Iran instead of short promoter in pQE vector to evaluate size effect between both promoters. We found no difference at EGFP intensity run by two different size promoters under the fluorescence microscope. Beyond the morphological plasticity, the EGFP is believed to be a non-toxic biological molecule (11, 13), but itfs over expression in a few instances is shown to be cytotoxic in transgenic mice even to cause dilated cardiomyopathy (14). Whereas the combination of these sequences in both our designed constructs successfully showed the faint and normal expression of EGFP in mouse pre-implantation stage embryos. It was regarded to specific data in which the faint-moderate expression of EGFP in transgenic embryos is compatible with normal embryogenesis in mice (15). The expression of EGFP protein into the cytosol and itfs solubility were not seemed to be toxic because of normal and high quality production of embryos. The green fluorescence observed in the morulae stage grew faint in all of the blastocyst stage embryos.We performed to select embryos at the blastocyst stage. This prevent firstly the effect of transient expression, result of the epichromosomal, and non-integrated transgene and secondly mosaic expression that may be due to the integration of the transgene after the first round of the DNA replication and thirdly for observation of GFP intensity in embryos (16, 17). In both mice and human, 15-50 % of embryos die during the preimplantation period from mechanisms that are largely unknown thus the low rate of surviving EGFP positive embryos comparing to only medium and physical treatments could be partly from the physical damage caused by microinjection and gene integration(Table I). We however decrease this matter because T10E0.1 buffer injected eggs showed clearly higher percentage of blastocyst development comparing to EGFP injected embryos. On the other hand, application of IVF wasdone firstly because of controlling both transgenic and non-transgenic embryos affected by medium and secondly out-coming sperm mediated gene transfer to provide some study about sex selection in livestock industry. It is related to accelerate genetically additive value of elite animals using blastomer biopsy and followed embryo sexing by PCR analysis (18, 19). It is interesting referring the data from Devgan et al (2004) that the developed blastocyst expressing EGFP can develop in post-implantation stage as well as reproductive out come of transgenic animals (14). As shown in the experiment by novel EGFP transgene, it is easy to classify transgenic eggs with high morphological plasticity before implantation stage with 100% of accuracy. Many mixed transgenic and non-transgenic embryos were separated under fluorescence microscope and were subjected to PCR analysis and the effect of new transgene was confirmed by standard grading method. Furthermore it was shown briefly the result of physical and transgene injected embryos that it needs to be large analysis on statistical basis. Conclusion The results of current mouse simulation model imply that an efficient production and propagation of transgenic livestock can be achieved by co-injection of every economical gene with this novel EGFP transgene and also it can be suitable gene cassette for numerous experiments and behaviors of proteins in living cells. Achnowledgments We are grateful to all the staff of Research and ClinicalCenter for Infertility for the use of the transgenic facility and Dr. Hiroshi Yasue (Genome Research Department, National Institute of Agro-biological Sciences, Tsukuba, Japan) for his invaluable critical reading of the manuscript. This study was supported by grants from National Network for Molecular Medicine, Pasteur Institute of Iran. References
© Copyright 2007 - Iranian Journal of Reproductive Medicine The following images related to this document are available:Photo images[rm07030f6.jpg] [rm07030f1.jpg] [rm07030f2.jpg] [rm07030f4.jpg] [rm07030f5.jpg] [rm07030f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}