 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Iranian Journal of Reproductive Medicine Vol. 6, No. 2, Spring, 2008, pp. 57-64 Cytogenetic and molecular genetic analysis of dicentric Y chromosome and its relation to male azoospermia Mir Davood Omrani1* Ph.D., Javad Karimzad Hagh2* Ph.D., Wolfrom Klein 3 M.D., Jurgen Gebauer 2 Ph.D. 1Department of
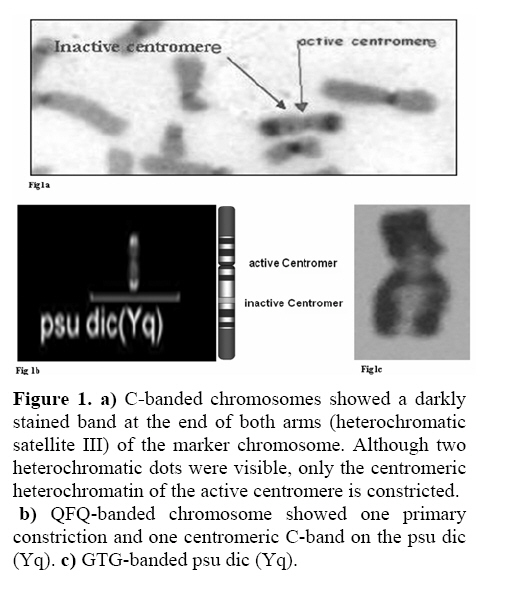
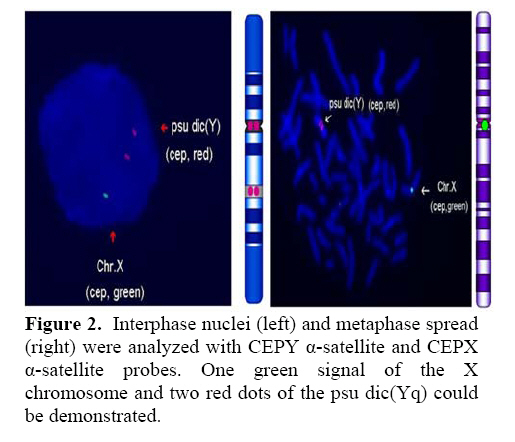
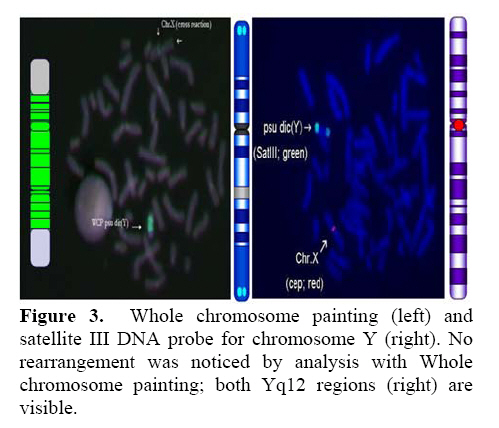
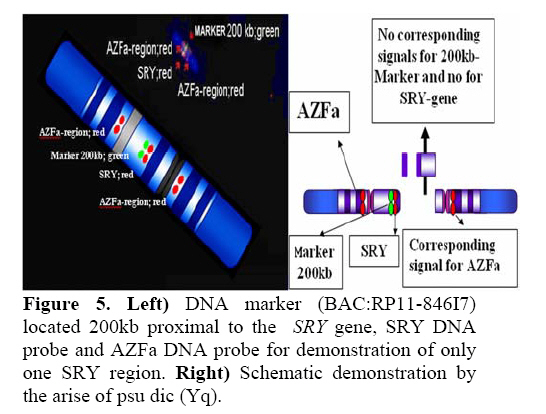
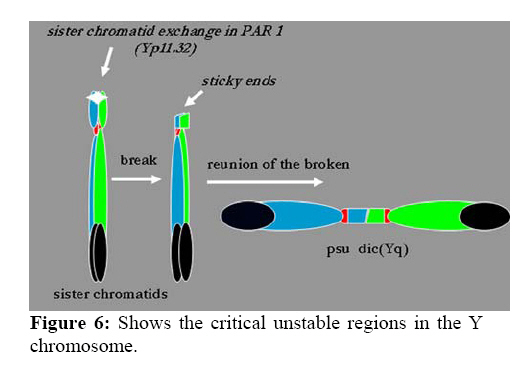
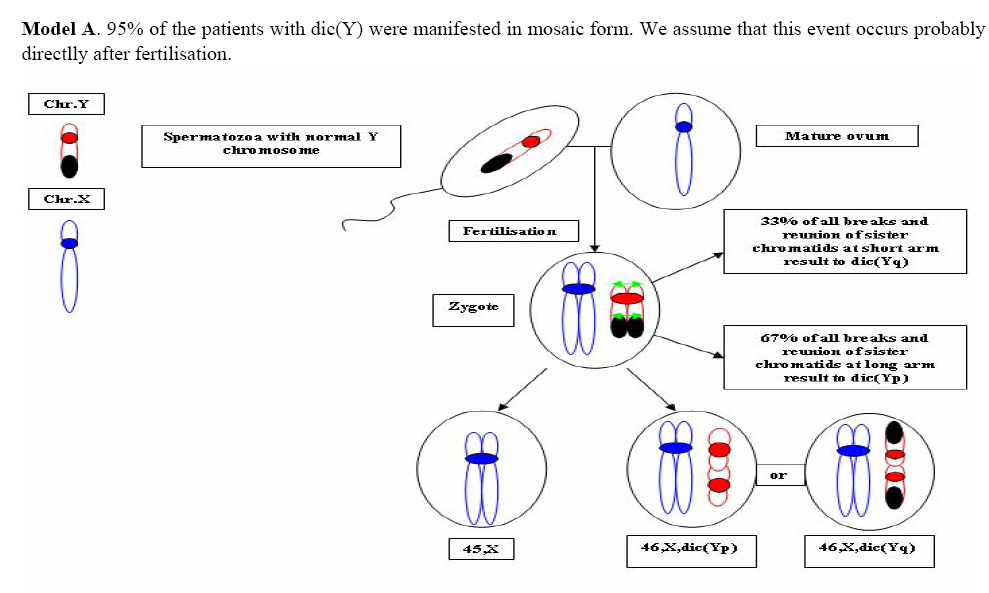
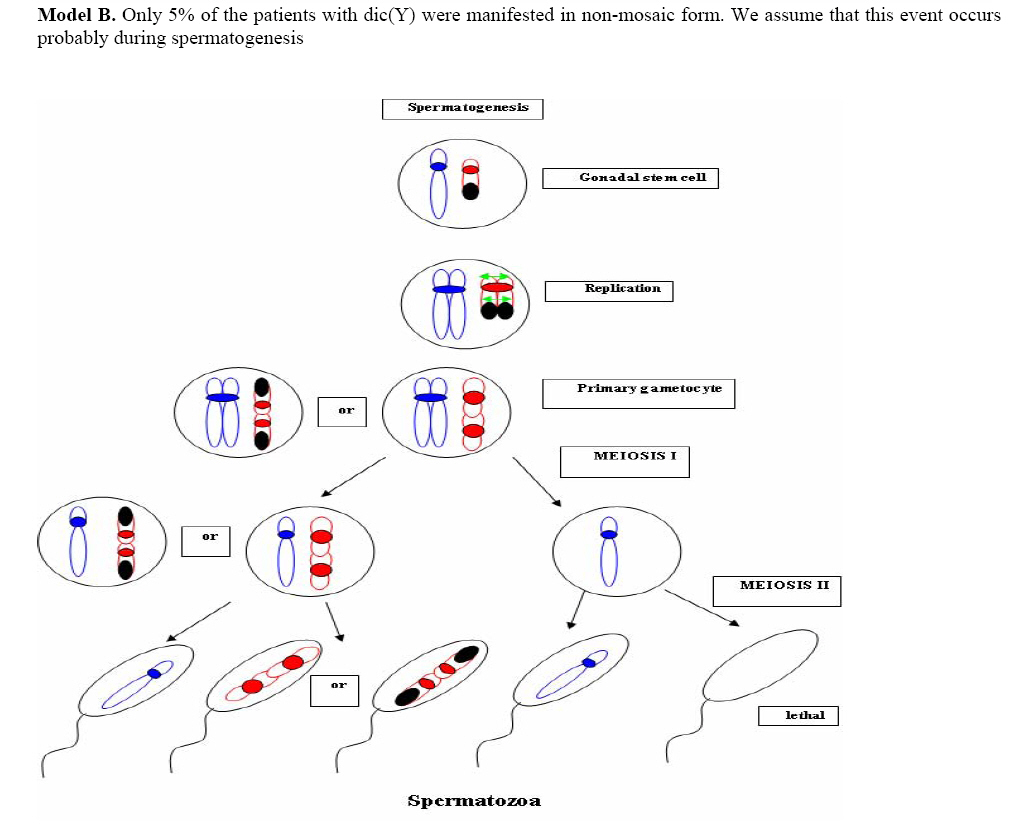
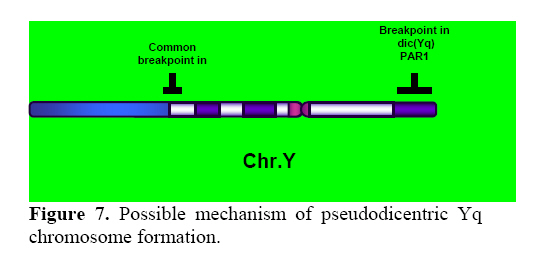
Genetics, Urmia Medical Sciences University, Iran. Received: 19 January 2008; accepted: 29 May 2008 Code Number: rm08010 Abstract Background: Cytogenetic analysis, Y-chromosome microdeletion screening, FISH techniques and other genetic methods have helped in finding the causes of infertility in azoospermic or severe oligoazoospermic cases in the last decade. Key words: Dicentric Y chromosome, Dic (Yq), FISH, Mosaicism, Azoospermia. Introduction Male infertility depends on several non-genetic and genetic conditions. Non genetic risk factors are; testicular maldescence,structural abnormalities of the male genital tract , varicoceles , chronic illness, genital infections, medication and exposure to chemicals. Idiopathic oligozoospermia or azoospermia in men could be due to deletions of the azoospermic factor region (AZF) of the Y-chromosome (1), mutations in the cystic fibrosis transmembrane conductance regulator (CFTR) gene (2) or numerical and structural aberrations of autosomes or the Y chromosome. Y chromosome inversions that dislocate the centromere close to the heterochromatin and satellites on Yq do not have clinical consequences. Structural aberrations of the Y chromosome, however such as deletions, rings, isochromosomes, dicentrics and Y-autosomal or X-Y translocations can affect the phenotype and are often associated with male infertility. The short arm of the Y chromosome is known to play an essential role in testis differentiation (3), whereas the azoospermic factor (AZF) genes, responsible for spermatogenesis are located on the distal region (interval 6) of the long arm (4). Deletions of this region have also been observed in 13% of azoospermic males with cytogenetically normal Y chromosomes (5, 6). Recently, two genes, RBM1 (RNA-binding motif 1) and DAZ (deleted in azoospermia), have been shown to map to a small region of the long arm of the human Y chromosome which is deleted in azoospermic males (7). Among structural abnormalities of the Y-chromosome, dicentric Y chromosomes were found in 26.5% of 468 patients by Hsu et al. (8). The incidence of abnormal karyotypes with dicentric Y chromosomes was 0.3% among 1523 azoospermic males examined by Yoshida et al. (9). These isodicentric Y chromosomes correspond to two different types: idic (Yq), refers to a dicentric Y chromosome with two complete long arms (Yq) and two symmetric but incomplete portions of the short arms with breakpoints occurring in the short arm of the Y-chromosome (Yp); in a idic (Yp) chromosome the Yq arms fuse with breakpoints at either Yq11 or, a more rarely Yq12 and with two short arms (Yp) intact. The sites of breakage and fusion at Yp and Yq are variable (10). Most cases (95%) with a dicentric Y chromosome were mosaicisms (8). These mosaicisms may be more or less complex depending on its meiotic or post-zygotic origin and on its mitotic stability, and usually includes a 45,X cell line (11). According to the structure of the dicentric Y chromosome, the Yp and Yq breakpoints, and the types of mosaicism, patients show a wide range of somatic, genital, and gonadal phenotypic manifestations (12). In a review by Hsu et al. (8) it was shown that more than 40.9% of subjects with dicentric Yq chromosome were phenotypicaly females, 31.8% were phenotypicaly males, and 27.3% had different degrees of intersexuality. It was possible to use PCR methods to determine the presence or absence of specific Y chromosome sequences but the obtained results do not provide enough information to analyse the structure of abnormal Y chromosomes (12). Therefore many investigators use other complementary method such as DNA probes for specifying Y chromosome sequences (12). In the present study, we analysed the structure of a dicentric Yq pesudochromosome and its breakpoints in an azoospermic male by conventional cytogenetic, molecular cytogenetic such as FISH and molecular genetics techniques in order to identify the causes of his infertility. Materials and methods The patient examined was a 38-year-old Iranian male ascertained of infertility. He has four healthy sisters. His height was 156 cm and his weight was 68.0 kg. His testes were very small and hard. Semen analysis showed azoospermia and the semen volume was 3.5 ml. Serum follicle stimulating hormone (FSH), luteinizing hormone (LH), testosterone and prolactin values were in normal ranges 7.7 U/L, 3.9 U/L, 5.2 ng/ml and 9.6 ng/ml respectively. Thyroid hormone tests were within normal limits. Pathocytologic examination of a testis biopsy revealed a large number of Leydig cells and seminiferous tubules but total absence of germ cells. Basement membrane thickening was partially seen. Cytogenetic analysis of peripheral blood lymphocytes was performed according to standard protocols using Giemsa banding (GTG), Quinacrine banding (QFQ) and Centromer banding (CBG) (13). In addition to conventional karyotype procedure, a high resolution chromosome analysis technique using thymidin for synchronization purpose was also carried out. Fluorescence in situ hybridization (FISH) was performed with whole chromosome painting (WCP Y; Q-Biogene) for Chromosome Y , FISH analysis of the Y and X chromosome α-satellite DNA probes of the centromic region, satellite III DNA probe for chromosome Y (Q-Biogene) and SRY-DNA-probe (Vysis). Further FISH analyses were carried out on metaphase spreads or on interphase nuclei using BAC probes. After culturing and DNA preparation, BAC-DNA was completely sonicated and labelled using the universal linkage system (ULS) (Q-Biogene). BAC probes were directly labelled with either, dGreen (green signal) or Rhodamin (red signal), according to manufacturer’s instruction. Subtelomeric Xp,Yp and Xq,Yp regions were screened using BAC probes: RP11-800K15 and Rp11-479B17 respectively. RP11-800K15 BAC probe also covers the SHOX gene, and it was used for detection of deletion in the SHOX gene in Yp and Xp regions. BAC: RP11-846I7 which is located 200kb proximal from SRY gene was applied to analyze deletion of SRY genes in the pseudodicentric Y chromosome. The AZF region was also analyzed with FISH probes as follows: AZFa region with BAC: RP11-460B2, DBY-gene green, AZFb with BAC: RP11-450B24, RBM1Y-gene, red and AZFc with BAC: RP11-140H23, DAZ-gene, green. All hybridizations were carried out for 12-18 hr at 37°C following a standard protocol (13). Slides were evaluated using an Axioscope 2 plus (Zeiss, Germany) and the Ikaros and Isis software (Meta system, Germany). Polymerase chain reaction amplification of the three AZF loci Genomic DNA was obtained from peripheral leukocytes using the Nucleon Kit II (Scotlab, Wiesloch, Germany). A set of 20 Y-chromosome specific STSs spanning the euchromatic region of Yq from centromere to interval 7, with particular interest in interval 6 (the AZF region), was tested. In order to check the AZFa region PCR amplification using sY81, sY83 and sY121, STS were carried out. Using STSs sY128, sY130, sY133, and sY143, the AZFb region was analyzed. The AZFc region was screened using sY147, sY242, sY231, sY149, sY254 and sY255 STSs markers. In addition sY202, sY158 and sY157 were included corresponding to the downstream region of DAZ as well as sY14 for the SRY gene and sY274 for next of SRY region. Each PCR included a sample of female genomic DNA as a negative control. A sample was considered negative if a product of the expected size was not obtained after three PCR attempts. The PCR program was: amplification of DNA by 35 cycles with 94 °C for 50 s, 57 °C for 30 s and 72 °C for 90 s; including an initial denaturation step at 94 °C for 2 min and a final extension step at 72 °C for 10 min. The PCR products were separated on 1.5% agarose gels. Semen analysis was carried out using WHO criteria with a Nikon phase contrast microscope. Serum hormone concentrations of FSH, LH and testosterone were measured by solid-phase, two site chemiluminescent enzyme immunometric assay (Immulite; Diagnostic Products Corporation, Los Angeles, CA, USA). Normal ranges for men are FSH <10 mIU/ml; LH <10 mIU/ml; and testosterone 270–1070 ng/dL. Results Two distinguished cell lines 45,X and 46,X +mar were detected in G-band spreads (Figure 1c). By C banding, the abnormal marker chromosome showed a darkly stained band at the end of both arms (Figure 1a) indicating the presence of two distal heterochromatic regions. In general, only one primary constriction and one centromeric C-band were visible, and although in some cases two internal regions of centromeric heterochromatin were present, only one was constricted. QFQ- banding technique also confirmed the C-banding findings (Figure 1b). Interphase nuclei and metaphase spreads were also analyzed by FISH with CEPY α-satellite, CEPX α-satellite and non-α-satellite III probes. Results from 183 cells examined showed mosaicm between a monosome 45,X (51%) and a pseudodicentric Y chromosome (49%) cell line (Figure 2): mos 45,X/46,X, psu dic (Yq) (qter→p11.32:: p11.2→qter). FISH with whole chromosome painting (WCP) for Chromosome Y (Q-Biogene), and satellite III DNA probe for chromosome Y (Q-Biogene) did not reveal any deletion and chromosomal translocation (Figure 3). Subtelomeric Xp,Yp region screening as well as SHOX gene tracing in this region, using BAC probe: RP11-800K15 has shown that the subtelomer signals Yp and SHOX gene signal in this region were lost in psu dic (Y) in both Yp arms but this signal is present in Xp arm (Figure 4). The SHOX gene maps to the pseudoautosomal region 1 (PAR1) of the X and Y chromosome. Combined FISH analyses with a SRY-DNA probe, BAC: RP11-846I7 which is located 200kb proximal from SRY gene and AZFa region with BAC: RP11-460B2 as control showed only one SRY gene in the pseudodicentric Y-chromosome (Figure 5). Analyses of the AZFabc region with several FISH probes and 20 STS markers did not show any deletion. There is no large or partial deletion in the q-arm. Moreover no translocation between the Y-chromosome and other autosomal chromosomes was identified. Discussion Reviewing 600 published cases with Y chromosome aneuploidy revealed 468 (78%) of them had structural abnormalities in mosaic and non-mosaic form [Y/autosomal translocation (27.8%), dic (Yq and Yp) (26.5%), Yq-(19.2%), Y/X transloction (11.2%), r(Y)(7.2%), i(Yq)(2.6%), Yp-(2.3%), i(Yp)(1.7%) and Y/Y(1.5%) translocation]. Among these 468 cases, only 124 (26.5%) of the cases showed a cell line with dicentric Y chromosome in their cultured lymphocytes. The majority (67%) of the known dicentric Y chromosome have breakpoint in their long arm, resulting to dic (Yp) chromosome, compare to dic (Yq) chromosome with breakpoint in the short arm of Y chromosome. More than 95% of dic(Y) chromosome manifest in mosaic form (8, 10, 11, 25, 26, 33). The breakpoint in the dic (Yq) seems to be limited more in the PAR1 region (pseudo autosomal region 1), whereas the dic (Yp) appears to have a common breakpoint (70%) in distal Yq11 region where eurochromatin region begins (10, 11) (Figure 6). The acentric fragments were generally lost after fusion between two arms of the Y chromosomes. Different mechanisms for the formation of dicentric isochromosomes had been described (14-16). Based on these informations it is possible to draw two different Models A and B for the dic (Y) development pathway. The mechanism of sister chromatid breakage and reunion seems most evident (14-16). This mechanism of breakage in the short arm of a Y chromosome and reunion of the broken ends is presented in Figure 7. The phenotype of patients with dicentric Y chromosomes differs from Turner like females to infertile males. The structure of the dicentric Y isochromosome, the Y chromosome breakpoints and the level and tissue distribution of mosaicism are the variables thought to be responsible for the variable phenotypes. However for dicentric Y-chromosomes it is still difficult to predict phenotypes from karyotype findings. Of 50 cases reviewed with dic(Yq), 22(44%) cases were phenotypically female, 16(32%) were phenotypic male and 12(24%) had intersex features. More than 95% of the dic (Y) chromosome cases were in the mosaic form(20). Our patient had a mos 45, X/46, X, psu dic (Yq) (qter→p11.32::p11.2→qter) mosaicism. The normal differentiation of the testis (apart from germ cells) in our patient showed that there had been SRY expression above the critical threshold to induce testis differentiation. FISH analyses with SRY-probes and the loss of one DNA marker 200 kb distant of the SRY-gene clearly showed that one SRY-gene is missing (only 50% SRY-gene-product). Unfortunately no information about mosaicism in other tissues of our patient is available, because we had no access to fibroblast cells or gonadal tissues. Some studies suggest that the reason for the presence of the mosaicism phenomenon in the dic (Y) chromosome patients were mistakes occurring during gametogenesis before the spermatid-stage, but since the majority of these patients show different proportion of mosaic cell lines in their tissues, therefore it is possible to assume that such occurrence in mosaic cases is most likely an event in early division stage after fertilisation (a postzygotic event). The double presence of the Y-chromosome genes in some tissues, in which Y-genes have high expression, is probably a disadvantage for the cell proliferation and differentiation. Therefore it is possible that the cell line without dic(Y) can divide faster compare to cell line with dic(Y). Review of the published mosaic cases’s data support this hypothesis. Also finding an extremely increasing rate of 45, X cell line in the fibroblast and female gonadal tissues support this opinion According to the FISH results described in Figure 4 the subtelomeric regions in both Yp arms of the dicentric Y chromosome must be lost. The BAC probe RP11-800K15 used for subtelomeric analysis covers the SHOX gene too. The Short Stature Homeobox-containing gene, (SHOX), located on the distal ends of the X and Y chromosomes, encodes a homeodomain transcription factor responsible for a significant proportion of long-bone growth. Patients with mutations or deletions of the SHOX gene, including those with Turner syndrome [TS] who are haploinsufficient for SHOX, have variable degrees of growth impairment, with or without a spectrum of skeletal anomalies consistent with dyschondrosteosis (17). It is already known that the distal ends of the short arms of the X and the Y chromosome are identical composed of 2.6 Mb DNA sequences (18). This region is called pseudoautosomal region 1 of the short arm (PAR1). Because Xp terminal deletions invariably result in short stature irrespective of the breakpoints, and small Yp terminal deletions equally lead to short stature, it has been suggested that a growth gene escaping X-inactivation resides in the PAR1, whose loss causes short stature in both sexes (19). Our case is the first report with 45, X/46, X, dic (Yq) karyotype which has been screened for SHOX gene using FISH analyses with evidence both SHOX genes missing at the psu dic (Yq). The SHOX gene maps to PAR1-region on the short arm of Chromosome X and Y. A reduced copy number of this gene in 332 adult turner patients with non-mosaic 45,X karyotype led to 100% short stature, whereas 52 turner patients with non-mosaic 46,X,Xp- karyotype led only to 88% (47) short stature, suggesting its key role for normal development in females (20). Discrepant to the key role of SHOX genes is the finding of the 10 phenotypicaly female individuals with non-mosaic 46, X,Yp- karyotype who had all a normal stature (8), in comparison with a non-mosaic 46,X,Xp- karyotype (88% small stature) suggesting that the located genes in Yq have probably more important role for normal development compared to Xq in female individuals with respect of inactivation process of the X-chromosome. Of 15 phenotypicaly male patients (including our case) with mosaic 45,X/46,X, idic (Yq), 61% had short stature (20), suggestive for a common feature in this individuals. Of course, stature is a multifactorial trait influenced by environmental factors and multiple genes. One of these putative genes was named growth control Y, (GCY) (28, 29). Several cytogenetic studies have shown that deleted or morphologically altered long arm of the Y-chromosome is found in patients with short stature (29-32). Other study reviewed that 29.2% of 48 phenotypic males non-mosaic 46, X, del(Y) (q11) karyotype had short stature (8) which maybe confirm this hypothesis. Our patient has a borderline small stature (156cm). This borderline small stature is caused probably of 45, X cell line and a deletion of the SHOX gene in the dic (Yq) chromosome (with only 50% of gene product). When a dicentric chromosome arises in early embryos, one of the two centromeres is usually inactivated increasing the stability of the dicentric chromosome (8, 12, 21). In these dicentric chromosomes that act as monocentric chromosomes (20), only one primary constriction is visible, but a second C-band mark the position of the inactive centromere (pseudodicentric chromosome). In our case two centromeres could be demonstrated by CBG- and QFQ-banding, one constricted and one without constriction. It is suggested that affected people with dic(Y) have a risk for the development of gonadoblastoma. None (0%) of 15 (including our cases) studied phenotypic male with mosaic 45,X/46,X, idic(Yq), ~11% of 18 phenotypic female and ~16% of 12 phenotypic intersex developed gonadoblastoma whereas the 4 (36.6%) of 11 reported female patients with non-mosaic 46,X,Yp- and 16 (24.5%) of 65 reported female patients with mosaic 45,X/46,XY had been developed gonadoblastoma (20). These studies support probably the hypothesis that the female who have mosaic 45,X/46,XY or 45,X/46,X, dic(Yq) and also non-mosaic karyotype 46,XYp- should classified in groups with high risk of gonadoblastoma development. Our patient is azoospermic. From anamnesis data we did not find any non-genetic risk factors, which can explain azoospermia. In addition, we could not find any deletions in the AZF-region. Probably the azoospermia in our patient could be explained by the abnormal Y chromosome. In meiosis the X- and Y-chromosome of normal structure join to form an XY-body or sex vesicle by end-to-end pairing of the short arm pseudoautosomal regions (PAR 1). With the loss of the PAR 1 region in our pseudodicentic Y-chromosome no regular XY-body can be formed. Pairing of the X- and Y-chromosome appears to be necessary for the completion of meiosis and the formation of sperm (22). In literature only 12 adult males with idic (Yq) had been published; (8, 11, 23, 24, 34), which 9 (75%) of these men showed azoospermia, too. In addition to this consideration the 45, X cell line, which may affect the normal gonadal development, could be responsible for the azoospermia. When the pseudodicentric Y-chromosome of our case induces sterility, we must assume that this structural aberration had been occurred de novo. Acknowledgements We would like to thank Dr. Vera Kalscheuer, Max-Planck-Institute of Molecular Genetics (Berlin), for preparing BACs-DNA. We are also very grateful to Prof. Brigitte Royer-Pokora, Dr. Barbara Leube and Mr. Dipl.-Ing. Manfred Beier for their efforts in this work. Permission was obtained from Urmia Medical Sciences University ethical committee (permission number 20013/12/2000). Reference
© Copyright 2008 - Iranian Journal of Reproductive Medicine The following images related to this document are available:Photo images[rm08010f3.jpg] [rm08010f5.jpg] [rm08010f7.jpg] [rm08010f2.jpg] [rm08010f8.jpg] [rm08010f9.jpg] [rm08010f4.jpg] [rm08010f6.jpg] [rm08010f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}