 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Iranian Journal of Reproductive Medicine Vol. 7, No. 2, Spring, 2009, pp. 59-64 The effects of diazinon on testosterone, FSH and LH levels and testicular tissue in mice Esmail Fattahi1 Ph.D., Kazem Parivar2 Ph.D., Seyed Gholam Ali Jorsaraei3 Ph.D., Ali Akbar Moghadamnia4 Ph.D. 1Department of Biology, Ayatollah
Amoli Branch, Islamic Azad University, Amol, Iran. Received: 23 October 2008; accepted: 3 May 2009 Code Number: rm09011 Abstract Background: Diazinon (DZN) is an organophosphate insecticide which
is used worldwide in agriculture. The
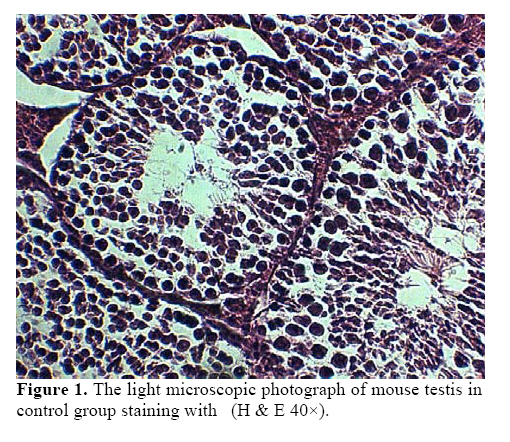
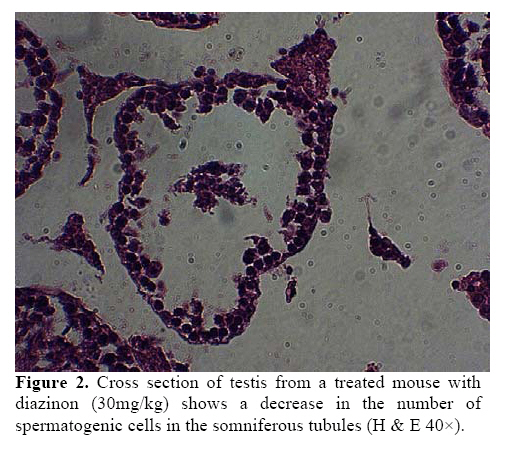
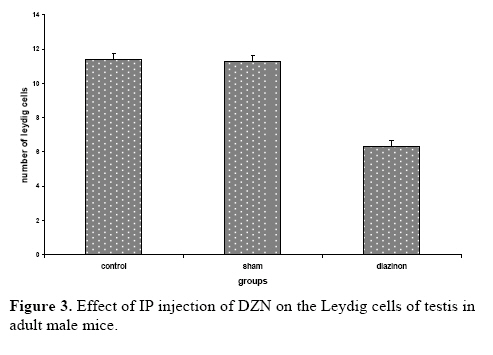
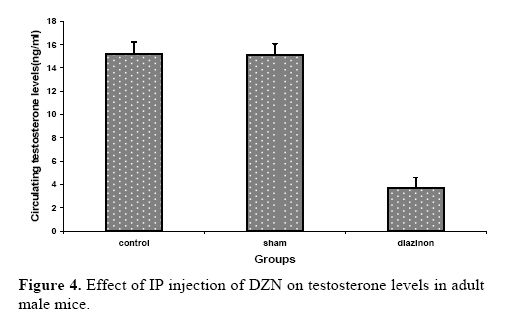
exposure to this chemical might lead to damages to the living systems. Key words:Diazinon, Leydig cells, Testosterone, LH, FSH. Introduction Diazinon (O, O-diethyl O - 2-isopropyl –6- methyl pyrimidinyl -4- g- 1- phosphorothioate) is insecticides widely used in agriculture and pest control in the environment, which can be highly toxic (1,2). “DZN is bioactived by cytochrome P450 enzymes through desulphuration to its corresponding Oxon derivative” (3). “These compounds exert inhibition on cholinesterase (ChE) activity by phosphorylation the catalytic site of the enzymes” (4). These are associated with male reproductive system, including chromatin alterations in sperm nucleus. The main toxic action of organophosphates (OP) is phosphorylation of proteins. Chemical changes in sperm nuclear proteins (protamines), which pack DNA during the last steps of spermatogenesis, is part of the cause to male reproductive toxicity (5, 6). Some studies showed that malathion and diazinon cause significant decrease in: oocytes survival, “regulation of genes encoding proteins involved in transcription (BP75), translation (ribosomal protein S5), and mitochondrial function (cytochrome oxidase subunits I and III), therefore providing evidence for OP insecticides as toxicants for mammals oocytes during the early oogenesis” (7). The insecticides that used in commercial formulation could be toxic and harmful to the developing embryo and in-vitro fertilization (8). “OP agrochemicals frequently alter human health or development, typically because they have endocrine agonist or antagonist activities and alter hormone-regulation of gene expression” (9). So, many adverse effects by DZN have been described so far including alterations in blood factors (RBC, Hb and Hct), plasma levels of testosterone and blood glucose (10). “Toxicity of OP mainly results in neurotoxicity due to its oxygen analogues (oxons), formed during the OP oxidative activation. OP alters semen quality and sperm chromatin and DNA at different stages of spermatogenesis. Oxons are more toxic than the parent compounds” (11). These compounds can influence the testes, for example decreasing the testis weight, sperm motility, grading and sperm viability, also they increase the sperm abnormal forms (12-13). Furthermore the OP overexposure through cutaneous absorption and inhalation has followed by neurological, skeletal and endocrine effects (14). Diazinon leading to the induction of lipid peroxidation, altrations the activities of antioxidant enzymes and DNA damage using in vitro (15). “Sublethal doses may lead to reduced growth and reproduction aquatic invertebrate, interference with algae-invertebrate interactions, reduced egg-production, delayed sexual maturity, adverse behavioral modifications in mammals and in fishes affect reproductive behavior and disrupt ovarian structure” (16,17). Some studies showed that exposure to environmental chemicals such as diazinon and malathion result in cancers or reduced reproductive functions in animals and human (18). However, there is little information about toxicological effects of pesticides, including diazinon, on the structure of testis and sex hormones in the male mice. Therefore, the main objective of this study was to investigate the effects of diazinon on the reproduction organs of mice. Materials and methods Chemicals Diazinon (DZN), a commercial formulation (60% active ingredient), was obtained from Shimi Keshavarz company of IRAN. The DZN was dissolved in corn oil and administered (i.p.) in sublethal dose. Animals For this experiment, adult male Balb/C mice (10-12 weeks old), were obtained from Pastor Institute. Mice were housed in a separate room cages, under controlled conditions of temperature (22±3°C), humidity (50± 5%) and light (12 light: 12 dark cycle), with access to the powdered diet and to deionized water. The experiments were conducted at the Babol University of Medical Sciences. For this study, the male mice were divided into three groups. In the experimental group, DZN was injected to the mice as consecutive doses (30mg/kg i.p, five consecutive days per week for one month). Sham received only the vehicle under the same conditions (corn oil injection) and control group kept with no injection till end of experiment. Animals were sacrificed by ether, 7 days after the last DZN injection. Histopathological examinations After the last DZN injection and at the time of tissue collection, testes were immediately fixed in 10% formalin solution. For each testis, serial sections (5 µm thick) were made. These sections were stained by hematoxylin-eosin (H & E) and morphometrically examined by light microscopy. Morphometrical analysis of the testis Testes were examined for structural changes. The following parameters were recorded: weight and diameter of testis, numbers of germ cells, spermatocyts, spermatids, Leydig cells, and the seminiferous tubules diameter (STD) which was measured by light microscope equipped by eyepiece graticule. Hormones measurements Serum LH and FSH were measured by double antibody radioimmunoassay as previously described (19, 20). Serum testosterone was assayed using a competitive chemiluminescent immunoassay kit (DRG Co, Germany; and Pishtaz Teb, Iran). Statistical analysis Data were represented as means±S.E. The differences were compared for statistical significance by one way ANOVA followed by Tukey post hoc test. Differences was considered significant at p<0.05. Results Effects of diazinon on testes Normal spermatogenesis could be seen in the testes sections of the control group (Figure 1). However, different degrees of degenerative changes in testes were observed in the DZN treated group (Figure 2). Sublethal dose of DZN in experimental group (30mg/kg) markedly reduced the diameter of seminiferous tubules (p<0.001) (Table I), which showed a variety of cellular damages including decrease in the number of germ cells of the tubules (p<0.001) (Table I). In this study, interaperitoneal (i.p.) administration of diazinon significantly reduced numbers of spermatocytes: control (38.5±1.1), DZN (35.02±0.8) and spermatids: control (12.04±0.23), DZN (9.38±0.41) (p<0.001) (Table I). Weight of testis was reduced in the mice receiving diazinon (7.32±0.32 mg), compared to the control group (8.28±0.29 mg) (p<0.05) (Table I). Testes relative weights were significantly increased by diazinon (30mg/kg,i.p.) compared to the control group. There were significant differences between the number of Leydig cells in the control and DZN groups (11.37±0.36 and 6.29±0.36 in control and DZN groups respectively) (Figure 3). The mean numbers of Sertoli cells on the control group was 2.08±0.18 while in DZN group it was significantly reduced to 1.10±0.093 (p<0.001) (Table I). Macroscopic studies showed that diazinon reduced testis diameter in the consecutive doses of DZN (6.44±0.18) compared with control group (7.01±0.48) (p<0.001) (Table I). Effects of diazinon on sexual hormones Diazinon significantly decreased the serum testosterone levels (3.68±0.91) in the experimental group compared with control (15.2±1.01) (p<0.001) (Table I). The levels of serum FSH was significantly higher in DZN group (5.4± 1.05) compared with the control group (2.5±0.53) (p<0.001) (Table I). The LH levels of the control group and DZN group were (3±0.53) and (4±0.64) respectively which were significantly different (p<0.001) (Table I). Table I. The effect of IP injection of diazinon on testicular tissueand sex hormones.
Discussion Some studies have showed that DZN was capable of inducing structural and functional changes (18), and some biochemical alterations in the ovaries and testes (16). Our results demonstrate that administration of sublethal dose of diazinon can adversely effects in reproductive function by decreasing the mass of testis and testosterone blood levels, but increasing the concentrations of circulating gonadotropins. Some histopathological studies in animals have showed that diazinon treatment may inhibit spermatogenesis and make destruction of all kind of seminiferous tubules. Maintaining LH serum levels is very important for initiating and supporting spermatogenesis, hence degeneration of Sertoli cells and germinal cells may be due to high concentrations of circulating LH (21,22). OP cause microtubule disruption of epithelium (23), and finally leads to tubular atrophy (24) Also, spermatogenesis disruption might be due to increases in the serum LH concentration which is detrimental to the germinal cells (having a very important role in the first steps of spermatogenesis process) (25, 26). The diameters of germinal cells and spermatozoa have showed a significantly reduction after DZN treatment (16). We also observed a decreased in the number of germinal cells. This might be the cause of decreases in the number of spermatocyts and spermatids, which would eventually result in a decrease of spermatozoa. Finally DZN could reduce sperm production. Toxic effects of diazinon might be mediated by stimulation of lipid peroxidation, changes in the actions of antioxidant enzymes, DNA damage, and free radicals which involved in cell death (26, 27). Similarly we observed that DZN decrease the number of all cell types in mice testis. Several studies have showed increased frequencies of sperm aneuploidy observed in individuals occupationally exposed to DZN (28). Also several OPs have been extensively studied as genotoxic chemicals in mammalian germinal cells and spermatids (29, 30). Prolonged IP exposure to the DZN, decreased the body weight in consensus with the earlier reports, and that effect was due to diminished food and water intake in treated mice of DZN group (31). Decrease in testis weight revealed the fact that significant gonadotoxicity could be seen after prolonged exposure to DZN. Similar effects has been observed in many other studies after treating with low doses of Diazinon (16), which may be due to decreased acetylcholinesterase level and spermatogenic cells, sertoli cells and Leydig cells losses and testicular atrophy (32-34). DZN has also been described as genotoxic , mutagenic carcinogenic agents mutagenic agent using in vivo and in vitro models (35, 36), probably by sharing the alkylation properties of many other OP compounds (29). DZN exposure also has caused reduction in the Leydig cells (16). Our results demonstrated that the number of Leydig cells was decreased. This could be the cause of testosterone shortage, which is needed along with the Sertoli cells to carry out spermatogenesis (16). “Testosterone is required for differentiation of sex organs and production of sperms” (37). OP may decline serum steroid hormone levels by increasing steroid catabolism and elimination or directly inhibit steroid hormone production(38). In addition OP insecticide inhibits steroidogenesis in adrenal cells (39). Therefore,maintenance of testosterone levels very critically for spermatogenesis and fertility (40).(40), reduce serum testosterone levels due to effects of diazinon to might cause reduction in spermatogenesis and fertility in animals. “There are several possible mechanisms for the antigonadal actions of organophosphates; they may exert a direct inhibitory action on the testis; they may affect the pituitary, causing changes in gonadotrophins concentrations and thus subsequent spermatogenic impairment; or they may change the concentration of neurotransmitter” (41). “Antiandrogens can disrupt male differentiation by several mechanisms, including antagonism of receptor binding, or by inhibition of the production, transport, or metabolism of androgens” (39). The damage may have occurred by direct toxic effects of DZN on cells or tissue, but it might also occur because of imbalanced hormones levels. DZN may affect directly upon testis tissue or by entering into the pituitary gland which could cause changes of gonadotropins arise. In these actions of DZN, serum LH and FSH levels increased probably due to suppression of feed-back inhibition of anterior pituitary. The suppression of feed back inhibition may secondarily increase the LH and FSH secretion. The results of the present study indicate that DZN has a direct effect on pituitary, which lead to increase in circulating LH levels. Diazinon increase serum FSH levels after injection in mice. FSH has important effects on sertoli cells. Inhibin and other factors secreted by Sertoli cells cause increase of circulating FSH levels by feed back on pituitary. Increase of LH levels has toxic effects on testis tissue. In summary, diazinon has adversely affected the reproduction organ in male mice. The present results indicate that exposure to DZN has direct effects on mice testis and the imbalance of circulating testosterone and gonadotropins may reduce fertility. Acknowledgment This project was supported by a grant from Islamic Azad University, Ayatollah Amoli Branch. References
© Copyright 2009 - Iranian Journal of Reproductive Medicine The following images related to this document are available:Photo images[rm09011f1.jpg] [rm09011f4.jpg] [rm09011f2.jpg] [rm09011t1.jpg] [rm09011f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}