 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Iranian Journal of Reproductive Medicine Vol. 8, No. 1, Winter, 2010, pp. 1-9 Effects of vitamin E on ovarian tissue of rats following treatment with p-nonylphenol: A stereological study Malek Soleimani Mehranjani1 Ph.D., Ali Noorafshan2 Ph.D., Ahmad Hamta1 Ph.D., Hamid Reza Momeni1Ph.D., Mohammad Hussein Abnosi1 Ph.D., Monireh Mahmoodi1 M.Sc., Morteza Anvari3,4 Ph.D., Maryam Hazaveh1 M.Sc. 1Department of Biology, Faculty of Sciences, Arak University of Medical
Sciences, Arak, Iran. Corresponding Author: Malek Soleimani Mehranjani, Department of Biology Faculty of Sciences, Arak University of Medical Sciences, Arak, Iran. E-mail: m-soleimani@araku.ac.ir Received: 23 July 2009; accepted: 13 January 2010 Code Number: rm10001 Abstract Background: Para-Nonylphenol (p-NP) is one of the environmental pollutants
which cause reproductive system disorders. Key words: Ovary, p-Nonylphenol, Rat, Stereology, Vitamin E. Introduction Para-Nonylphenol (p-NP) is one of the endocrine disrupting compounds, which has estrogenic effects and is able to impair the endocrine system ( 1 - 3 ) through diverse mechanisms, such as receptor-mediated enzyme inhibition (1) and mimics the estrogen or androgen action (1, 2, 4). This chemical is used in the preparation of lubricating oil additives, plasticizers and surface active agents and also has been found in polyvinyl chloride which is used in the food processing and packaging industries (1). Investigations have shown that the p-NP causes dysfunction in male and female reproductive system (5-7) specially during the early phases of animal development (1). In addition it also produces oxidative stress (3, 8) through generating free radicals and impairing estrous cycle as well as folliculogenesis (9). Some investigators have reported that the p-NP exposure have caused the reduction of body and ovary weight as well as reproductive impairment. Cunny et al, showed that the treatment of female rats with 139mg/kg body weight/day of p-NP for a period of 90 days caused significant reduction of body and ovary weight (10). Treatment of rats for 20 days with 50 and 100mg/kg body weight/day of p-NP caused imbalance in estrous cycle whereas 100mg/kg body weight/day showed a significant reduction in ovary weight (11). Chapin et al, showed a significant reduction of the number of antral and graafian follicles after treatment of the rats with 350mg/kg body weight/day of p-NP in F2 generation (12). In another investigation, treatment of pregnant female rats from the day 7 of pregnancy till the maturation of the pups with 400mg/kg body weight/day of p-NP, the F1 generation have lost the fertility potential (13). On the other hand, antioxidants such as vitamin E can prevent the adverse effects of oxidative stress by inhibiting reactive oxygen species (ROS) (14,15). Investigation showed that the co-administration of vitamin E with Methidathion and Organophosphate insecticides (16) prevents the harmful effects of these chemical in the reproductive organs. Considering the above, we aimed to investigate the preventive effects of vitamin E as a strong antioxidant on structural parameters of ovary during its development in the rats treated with p-NP, as an oxidative agent, using stereological methods. Materials and methods Animals and treatments Male and female Wistar rats with average weight of 200±10g were purchased from Pasteur Institute, Iran and kept in the animal house of Arak University under standard conditions of temperature, light and food. After mating, pregnant rats were divided into 4 groups (n=6): control, p-NP (250mg/kg/day) (Acros Company, New Jersey. USA), vitamin E (VE) (100mg/kg/day) (Aldrich Company, USA) and p-NP+VE. The treatment was carried out from the 7th day of pregnancy till 21st day of postnatal through sucking period (17-19). From this stage, female pups (F1 generation) were divided into the same groups similar to the mothers and the treatments were continued orally by gavage till maturation (120 day) (1). Considering the high viscosity of p-NP and vitamin E, corn oil was used as a carrier (5,20), with the concentration of 50mg p-NP/0.1ml corn oil (21), and similar vehicle was employed for vitamin E. Tissue preparation At the end of the treatments period, the rats were weighed, anesthetized by diethyl ether (Merck company, Germany) and their right ovary were taken out, weighed and fixed in 10% neutral buffered formalin. After tissue processing, the samples were blocked in cylindrical paraffin blocks. Stereological study The orientator method was used to obtain isotropic uniform random (IUR) sections (22, 23). For this purpose, the cylindrical paraffin blocks containing ovaries were randomly placed on the φ-clock which each half of it was divided into 9 equal parts. By choosing a random number from 1 to 9, an appropriate cut was made along the selected number. The block was then placed on the θ-clock, each half of it was divided into 9 unequal sine-weighted parts, along its cut surface on the 0-0 axis and then the random number was selected and the cut was made along the selected number. Consecutive 5 and 20 μm thick sections were prepared using a microtome and stained with Hematoxylin and Eosin (H&E) (Merck company, Germany) method. The volume of ovary, cortex, medulla and corpus luteum (mm3) To estimate the
mean total volume of ovary, the images of 5μm thick sections were transferred
on the working table using the micro-projector (NeoPromar Leitz Germany) with
the magnification of 80. Counting probe was randomly superimposed on the images,
then the points were counted and the total volume of the ovary was estimated
using the Cavalieri methods applying the following formula (22): The number of follicles To estimate the number of follicles, the optical disector method was used. The average of 12 sections was selected from 20μm thick sections using systematic random sampling. The sections were studied using the Olympus microscope (BX41TE model) with 100x magnification and the microcator (ND 221 B, Heidenhain, Germany) connected to a computer. The nuclei of follicular cells were sampled by an unbiased counting frame superimposed on the monitor. Identification of the stage of follicles was carried out based on the Mayer et al classification (24). Any nucleus that lied in the frame and did not touch the left and bottom lines of the frames was selected. The number density (Nv) of different types of follicles was estimated as: The volume of oocyte, follicular cells and their nuclei To estimate the volume of oocyte, follicular cells and their nuclei the nucleator method was applied. This is a method for estimating the number-weighted mean volume (νN ). An average of 12 sections from 20μm thick sections was randomly selected and the selected follicles were then studied using the Olympus microscope with 100x magnification. On the selected cells with an unbiased counting frame, the distance from the center of the nucleolus to the oocyte membrane was measured ( ln ) to estimate the oocyte volume. To estimate the volume of oocyte nucleus the distance from the center of the nucleolus to the nucleus membrane was measured. The same measurements were carried out for the follicular cells and their nuclei. The volume of each was calculated using the equation Zona pellucida thickness (μm) To estimate the mean thickness of zona pellucida (ZP), an average of 12 sections from 5μm thick sections was randomly selected and studied with 100x magnification. To identify measurement sites, the specific line grid (3 parallel lines) was randomly superimposed on the sampled fields. The ZP thickness was measured using the orthogonal intercept method, in brief by measuring the length of a line extended perpendicularly from the inner membrane to outer surface of ZP at each intercept of the line of the grid with zona membrane, and was considered as orthogonal intercept (oi). An average of 110 measurements was made to calculate the harmonic mean thickness using following formula (27): Harmonic mean thickness = 8π /3 × Harmonic mean of orthogonal intercepts, where harmonic mean = number of measurements / sum of the reciprocal of orthogonal intercepts lengths =
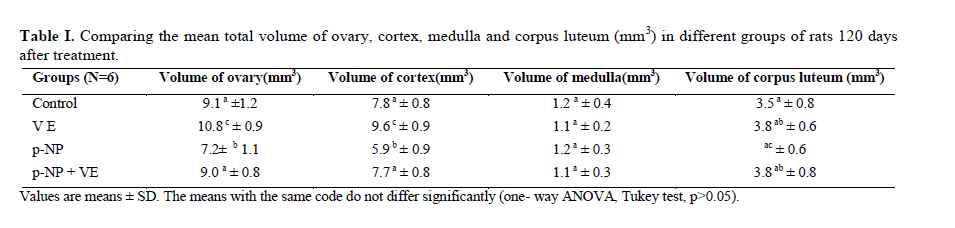
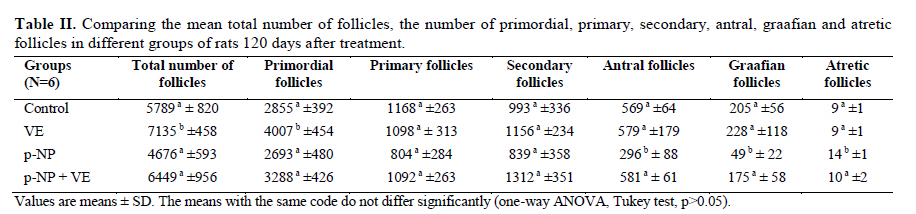
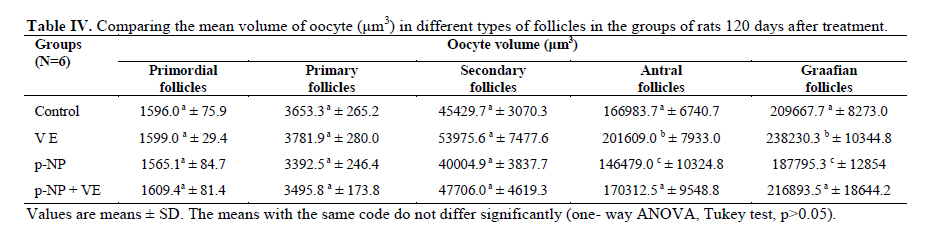
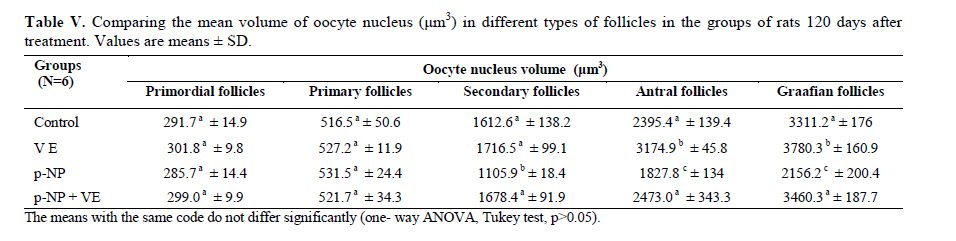
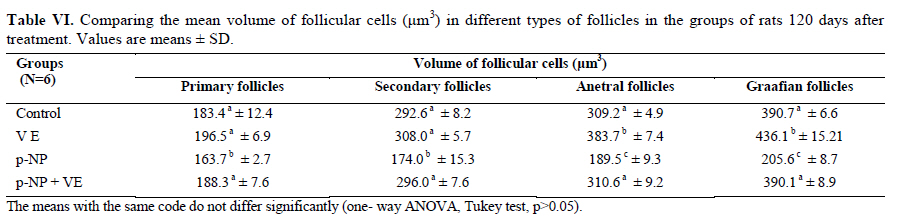
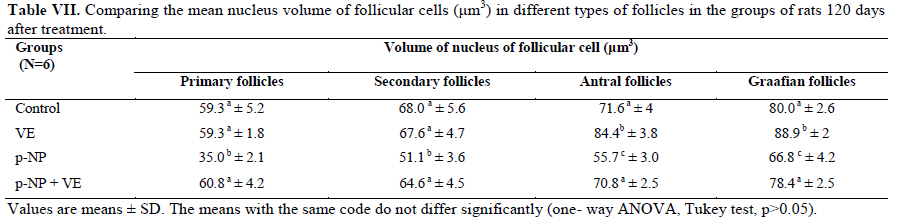
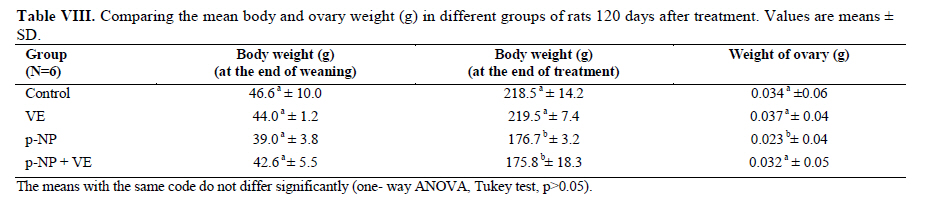
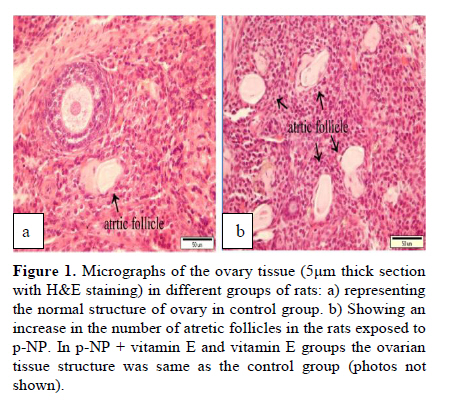
Statistical analysis The results were analyzed by one-way ANOVA, Tukey test, using the SPSS V11/0 software and the means were considered significantly different at p<0.05. Results The volume of ovary, cortex, medulla and corpus luteum The mean total volume of ovary and the volume of cortex showed a high significant reduction in p-NP group compared to the others (p<0.01), while co-administration of p-NP with vitamin E increased the volume of ovary and cortex to the control level. Comparing the mean volume of corpus luteum, a significant reduction was found in the p-NP group compared to vitamin E and p-NP + vitamin E groups (p<0.05) (Table I). The number of follicles The mean number of antral (p<0.04) and graafian follicles (p<0.04) reduced while the mean number of atretic follicles increased significantly (p<0.001) in p-NP group when compared to the other groups. In p-NP+vitamin E group, variations due to p-NP exposure were compensated to the normal level. In the group treated with only vitamin E, the mean total number of follicles (p<0.02) and the mean total number of primordial follicles (p<0.001) significantly increased compared to the other groups (Table II). The thickness of zona pellucida (μm) Comparing the mean thickness of zona pellucida in the secondary follicles (p<0.03), antral (p<0.001) and graafian (p<0.002) a significant reduction was found in the p-NP group, while simultaneous treatment of rats with p-NP + vitamin E compensated these reductions (p>0.05) to the level of control group. In addition, treatment with only vitamin E caused considerable increased in the zona pellucida thickness of secondary follicles (p<0.05) (Table III). The volume of oocyte (μm3 ) The mean volume of oocyte in antral and graafian follicles reduced significantly in p-NP group compared to other groups (p<0.04), while in the p-NP + vitamin E group, vitamin E compensated the reducing effect of p-NP to the normal level (p>0.05). The result also showed treatment with only vitamin E caused a considerable increase in oocyte volume of the mentioned follicles (p<0.001). The mean volume of oocyte in primordial, primary and secondary follicles showed no significant difference in any of groups (p>0.05) (Table IV). The volume of oocyte nucleus (μm3 ) The mean volume of the oocyte nucleus (μm3) in secondary, antral and graafian follicles showed a highly significant reduction in p-NP group compared to the other groups (p<0.001). Meanwhile the mean volume of oocyte nucleus in these follicles was increased highly significant following vitamin E treatment (p<0.001) in the p-NP + vitamin E group. In the vitamin E group, an increase in the volume of the oocyte nucleus (μm3) in antral and graafian follicles was seen (p<0.05). The volume of the oocyte nucleus of primordial and primary follicles showed no significant difference between the groups (p>0.05) (Table V). The volume of follicular cells (μm3) The mean volume of follicular cells in primary, secondary, antral and graafian follicles significantly decreased in p-NP group when compared to the others (p<0.001), while simultaneous treatment of vitamin E and p-NP compensated this reducing effect of p-NP in the above mentioned follicles (p<0.05). The treatment of rats with only vitamin E resulted in a significant increase in the mean volume of follicular cells in antral and graafian follicles (Table VI). The volume of nucleus of follicular cells (μm3) The mean volume of the follicular cells nucleus in primary, secondary, antral and graafian follicles showed a highly significant reduction in the p-NP group compared to the other groups (p<0.001). The simultaneous treatment of rats with vitamin E + p-NP compensated the reduction of volume of follicular cells nucleus due to p-NP in the above follicles. Meanwhile the treatment of rats with only vitamin E led to a highly significant increase in the mean volume of follicular cells nucleus in antral and graafian follicles (p<0.001) (Table VII). The body and ovary weight (g) The mean body weight showed a significant reduction in the p-NP and p-NP + vitamin E groups compared to the other groups at the end of treatment (p<0.01). In the p-NP group compared to the other groups, a highly significant reduction in ovary weight was observed (p<0.001), while in p-NP + vitamin E group, it was seen that treatment with vitamin E could compensate the reduction of ovary weight caused by p-NP (Table VIII). The histopathological findings An increase in the number of atretic follicles was found in p-NP group compared to the other groups. However, simultaneous treatment of p-NP and vitamin E seemed to normalize the number of atretic follicles (Figure 1). Discussion As the results showed a significant reduction in the weight of the ovary in the rats treated with p-NP was found which is in agreement with other investigations (28- 29) and also it is believed that this reduction is dose–dependent (10, 12). Atrophy of the ovary along with the lack of corpous luteum and reduction in the number of follicle could be considered as parameters involved in ovary weight reduction (20, 30). In addition, treatment with p-NP can reduce the level of gonadotrophins (29), known as an inhibiting factors of apoptosis in granulosa cells (31), therefore following p-NP treatment granulosa cells may undergo apoptosis and shrinkage which will finally lead to a reduction in ovary weight and volume. In another study the reduction in mitosis of granulosa and theca cells during folliculogenesis as a result of p-NP treatment was shown (32), which can affect the follicle growth and may also cause the reduction of ovary weight and volume. Co-administration of p-NP with vitamin E compensated the reduction in ovary weight and volume. This could be due to the fact that vitamin E can stimulate the secretion of Gonadotrophins(33, 34) which had been reduced as a consequence of p-NP. In addition it should be noted that vitamin E as a strong antioxidant (15, 16) may prevent weight and volume reduction in the ovary, due to the prevention of oxidative stress caused by p-NP (9). The results showed that the mean thickness of zona pellucida significantly reduced due to p-NP treatment. Follicular atresia and reduction in mitosis in granulosa and theca cells following treatment with p-NP has been reported by other investigators (1, 20, 32). Since it is believed that the follicular cells and oocytes are involved in production of zona pllucida (35) and in this study a significant reduction in the volume of oocyte nucleus, volume of follicular cells and its nucleus in secondary, antral and graafian follicles as well as the volume of oocyte in antral and graafian follicles following treatment with p-NP was found, therefore the reduction in the thickness of zona pellucida is expected. In addition, oxidative stress caused by p-NP (9) could be involved in the degradation of lipid and protein components of zona pellucida, as this effect has been reported on the chorion layer in zebra fish (36). As an outstanding result of this study, vitamin E not only compensated the reduction of zona pellucida thickness when given simultaneously with p-NP, but also normalized the reducing effects of p-NP on the other mentioned structural parameters. We also found that treatment with p-NP caused reduction in the number of antral and graafian follicles, which is in agreement with Chapin and co-workers, study (12). In the control and p-NP groups, the number of primordial, primary and secondary follicles were nearly identical, and a significant increase in the number of atretic follicles was observed in the p-NP group which is in agreement with other investigators (1, 20); therefore this increase is mainly relevant to the decrease of antral and graafian follicles, so it is concluded that p-NP effects are more obvious in the later stages of follicular development. On the other hand, it is reported that the increase in the number of atretic follicles is due to the disturbance of folliculogenesis and oogenesis as a result of oxidative stress and estrogenic property of p-NP which leads to a decrease in the number of follicles in preovulation stage (9, 20). In the present study vitamin E as strong antioxidant (15, 16) could prevent follicular degeneration and atresia as a consequence of p-NP treatment and also compensate the reduction in the number of follicles. The same results were also reported when vitamin E and vitamin C were given as antioxidant following treatment with methidathion, a substance which cause lipid peroxidation in the ovary and increase the number of atretic follicles (16). As mentioned above, in this study we also observed that the mean volume of oocyte and its nucleus in antral and graafian follicles decreased significantly in p-NP group. The same results were also obtained for the volume of follicular cells and their nucleus in primary, secondary, antral and graafian follicles. p-NP exposure can inhibit the secretion of follicle stimulating hormone (FSH) and luteinizing hormone (LH) which are vital for oocyte development (29, 37) and also causes apoptosis of granulosa cells which can be considered as another reason for FSH level reduction (31,38). Therefore, reduction in the level of these hormones retards the follicular phase and impairs oocyte maturation which leads to a decrease in the volume and the number of follicular cells. In addition, as a result of estrogenic property of p-NP (1), folliculogenesis impairment and retardation in oocyte maturation are expected as shown with other xenoestrogens (37, 39). Co-administration of vitamin E with p-NP could prevent the disturbance in the number of mentioned follicles caused by p-NP which is due to the antioxidant effect of vitamin E (15, 16) or the fact that vitamin E is able to stimulate FSH secretion(33, 34). The presented data also revealed that the mean body weight significantly reduced in p-NP group. Investigations have shown that p-NP as an environmental toxicant can reduce body weight through its estrogenic property which seems to be dose-dependent (1, 20, 40, 41). This could be due to the disturbance of hypothalamus-pituitary-ovary axis which leads to deficiency in growth hormone secretion as seen following the treatment of rats with octylphenol as another estrogenic substance (42). We also observed that p-NP caused anorexia and reduction in food consumption which can be another influencing parameter in weight loss as reported by other investigators (10, 40). Following treatment with p-NP an increase of T3 hormone level is also reported (29) which can be considered as another mechanism for reduction of body weight. However, treatment of rats with p-NP+ vitamin E as a strong antioxidant (14, 43) had no effect on weight reduction. Therefore, the weight reduction following treatment with p-NP may not be due to its oxidative stress or may be, the used dosage of vitamin E had not been sufficient. In conclusion, vitamin E could compensate the most adverse effects of p-NP treatment on ovary structure and also it was seen that the application of vitamin E only could increase the volume of ovary and cortex, the number of primary follicles and improved the main structural parameters of follicle in the late stages of follicular development. As the result of this study showed the consumption of vitamin E might be useful where ever the toxicity with p-NP is the matter of concern. Acknowledgment Authors would like to thanks Arak University, research and technology administration office, for financial support of this research. References
© Copyright 2010 - Iranian Journal of Reproductive Medicine The following images related to this document are available:Photo images[rm10001t3.jpg] [rm10001t8.jpg] [rm10001f1.jpg] [rm10001t1.jpg] [rm10001t4.jpg] [rm10001t5.jpg] [rm10001t7.jpg] [rm10001t2.jpg] [rm10001t6.jpg] |
| |||||||||

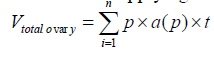 , in which
, in which 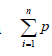 is the total number of points superimposed on the image, (t) is the thickness
of the section and a (p) is the area associated with each point. To obtain
the volume of the ovary compartments, the volume density for each was calculated
as:
is the total number of points superimposed on the image, (t) is the thickness
of the section and a (p) is the area associated with each point. To obtain
the volume of the ovary compartments, the volume density for each was calculated
as: 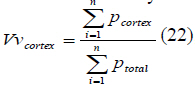 (22), where
(22), where 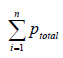 is the total number of counted points and
is the total number of counted points and 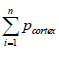 is the total number
of points superimposed on the cortex. The volume of cortex was then obtained
through multiplying the volume density (Vv) by the total volume of ovary.
is the total number
of points superimposed on the cortex. The volume of cortex was then obtained
through multiplying the volume density (Vv) by the total volume of ovary.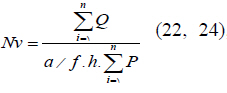 (22, 24), in which
(22, 24), in which 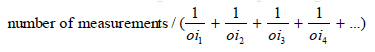
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}