 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
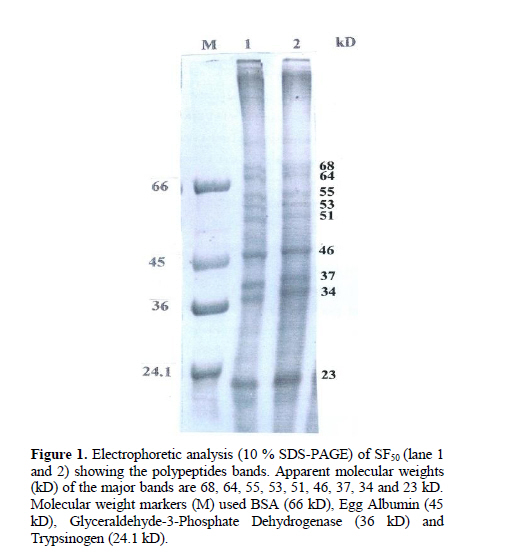
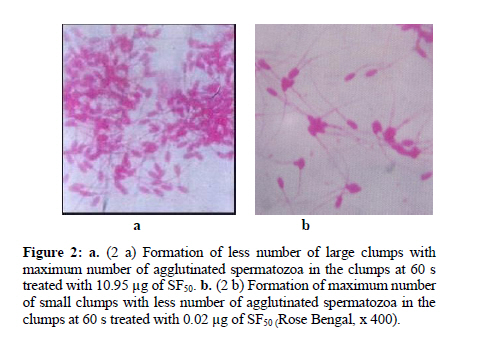
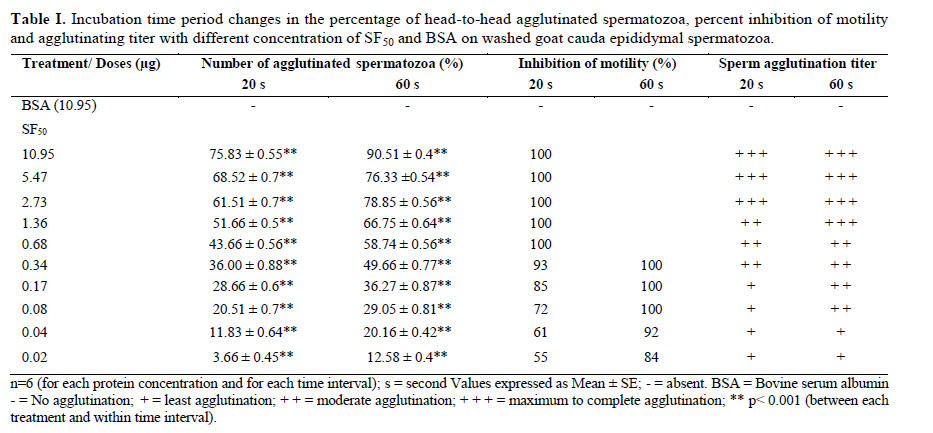
Iranian Journal of Reproductive Medicine Vol. 8, No. 1, Winter, 2010, pp. 10-17 A new sperm agglutinin factor from marine snail Telescopium telescopium: An evaluation with goat (Capra hircus) cauda epididymal spermatozoa Samata Maji M.V.Sc., Uttam Datta Ph.D., Manik Lal Hembram M.V.Sc. Department of Veterinary Gynaecology and Obstetrics, Faculty of Veterinary and Animal Sciences, West Bengal University of Animal and Fishery Sciences, West Bengal, India. Corresponding Author: Uttam Datta , Department of Veterinary Gynaecology and Obstetrics, Faculty of Veterinary and Animal Sciences, West Bengal University of Animal and Fishery Sciences, 37 and 68, Kshudiram bose sarani, Belgachia, Kolkata-700037 West Bengal, India. E-mail: uttamdtt@yahoo.com Received: 20 Jun 2009; accepted: 19 December 2009 Code Number: rm10002 Abstract Background: Lectins, the multivalent carbohydrate binding proteins are also employed to ascertain differentiation and characterization of the specific cell surface carbohydrate ligands/ receptors of different cell types including spermatozoa and posses novel biomedical importance. Key words: Agglutination, Lectin, Snail, Sperm. Introduction Mammalian spermatozoa emerging from the testis, display a highly organized and differentiated cell surface with membrane components arranged in discrete domains (1,2). The distinct cell surface topography of glycoprotein and other membrane components modify during passage through the male reproductive tract, plausibly play a crucial role in the physiology of spermatozoa during epididymal maturation (3-5), capacitation and acrosome reaction (6,7) as well as in sperm-egg interaction (8-10). For identification and understanding of the role of sperm surface components involve in maturation and fertilization, lectins (agglutinins) have been used to characterize such components (11- 13). Lectins, are multivalent high molecular weight glycoproteins with different sugar binding specificity, are able to agglutinate different cell types and bind specific carbohydrate moieties on the cell surface membranes particularly in terminal positions (14, 15). Lectins have been used to study/ characterize the carbohydrate-containing cell surface receptors in numerous biological systems and can be used to analyze variations in the distribution and density of exposed saccharides of the sperm plasmamembrane (SPM) during physiologic maturation and after ejaculation (16, 17). Lectin-binding has also been studied on spermatozoa of different species since glycoconjugates are believed to be functionally important in sperm-egg recognition and interaction. Hence, lectin-binding may therefore, bear some potential roles for biological in-vitro tests to predict functional activity of spermatozoa (18, 19). Importantly, agglutination assay may prove useful as a direct test for the occurrence of capacitation which could be used to develop/ compare surface properties of uncapacitated and capacitated mammalian sperm. Lectins have been isolated from variety of plants, animals and microbial sources (14, 20, 21). Invertebrates are also rich sources of different types of agglutinins (22) and are found in their haemolymph, albumin gland and in sex organs also. Agglutinins are glycoprotein in nature and play a fundamental role in the innate immune responses in invertebrates (14). Although various activities have been attributed to invertebrate lectins, however, reports on the activities / characters of agglutinin from Telescopium telescopium on the sperm cells are very scarce. In this study, a simple assay was developed to quantitate, SF50, a sialic acid specific lectin (23) on washed goat cauda epididymal spermatozoa, and at timed intervals during incubation, sperm samples were assessed for pattern and degree of SF50 induced agglutination, immobilization as well as surface alteration if any, in comparison with BSA treated spermatozoa as control. Materials and methods Collection of the snail and preparation of SF50 The cone snail Telescopium telescopium under phylum-mollusca, class-gastropoda, were collected from the estuaries of inter tidal zone during low tide at the Bay of Bengal near Sagar Island (22° 19′ N; 80° 03′ E) in West Bengal, India. 50 adult snails were collected randomly before the rainy season. The snails were kept in the laboratory at 28°C to 30°C in a tub containing sea water. Before the experiment, snails were allowed to empty their guts for a period of three days. The snails were kept under running tap water to clean the mud from outer surface of the shell and blotted to remove water prior to the experiment. Before being removed from their shells, snails were narcotized in isotonic (7.2%) magnesium chloride solution in a tray for 30 min. the shells were broken very carefully with a hammer. The whole body of each snail was removed and washed thoroughly with normal saline solution and blotted. Spermatheca gland from each snail was dissected out from the body of the snail. The dissected glands were minced carefully in a Petri dish, homogenized at 4°C with 4 volume of 0.15 M phosphate buffer saline (PBS; 8.76 g/L NaCl, 0.3 g/L NaH2PO4, 1.07 g/L Na2HPO4), PH 7.2 in a motor driven Teflon Dounch homogenizer The grinded tissue fragmented further by sonicator (6 µ amplitude) at 0°C with a Brownwill Biosonic IV, VWR sonicator set at 100% output for four 15 s brusts at ≥30 s interval for a total period of 1 hour. The sonicated material was spun in a Beckman SW28 swinging bucket rotor at 15,000g for 30 min at 4°C to discard the cellular debris. Cytosol fraction was aspirated carefully from the centrifuge tubes and ultracentrifuged again at 90,000g for 1 h in a Beckman L265B ultracentrifuge using a fixed angle rotor type 65 at 4°C. The tinged bluish coloured clear cytosol fraction was aspirated carefully by Pasteur pipette and kept in a sterilized glass beaker. The cytosol fraction was passed through membrane filter (150µ pore size) and transferred in a beaker with magnetic stirrer. Ammonium sulfate solution, 50%, was added drop by drop and allowed to stir overnight at 4°C. The solution was centrifuged again at 10,000 g for 30 min at 4°C. Precipitated protein pellet was collected carefully and dissolved in 2 ml of PBS. Dissolved protein solution was transferred into a dialysis bag (450 Å poracity) and dialyzed against 6 changes of PBS for overnight at 4°C. Dialyzed solution then transferred into sterile glass vials, lyophilized and kept at -20°C till experimentation. The lyophilized fraction was named SF50 (24). Estimation of total protein and PH of SF50 Total protein concentration of SF50 (1 mg/ ml in distilled water) was estimated (25) against standard solutions of BSA (globulin free). The readings were recorded by a spectrophotometer at 280 nm. The PH value of SF50 solution was estimated by a PH meter. Polyacrylamide gel electrophoresis (SDS-PAGE) of SF50 SDS-PAGE (10%) was performed (26) under reducing condition to identify the individual proteins from prepared SF50 solution (1 mg/ml in double distilled water). Approximately 40 µg of protein was loaded in each lane. Molecular weight markers (Bio-Rad) were also run parallely. After electrophoresis, the separating gel was stained with periodic acid-Schiff stain (27). Isolation and preparation of goat cauda epididymal spermatozoa Testes from two Black Bengal bucks were collected from local abattoir immediately after slaughter and were brought to laboratory in thermos flask containing normal saline solution (NSS; 0.9% ) at 37°C. Tunica albuginea were removed from the testes and washed thoroughly with NSS. Fat-pad, blood vessels, adipose and connective tissues were removed carefully. Whole epididymides were dissected out from each testis. As per anatomical position each cauda epididymis was cut and placed into a polystyrene Petri dish containing 5 ml of PBS. Cauda epididymides were minced carefully and allowed to suspend in the medium for 10 min. To obtain sperms, gentle pressures on the excised tissues were given by clean glass rod. The resultant suspension in the medium was filtered through nitex membrane to free the cellular debris. The filtrate was collected into a clean glass test tube, centrifuged at 500 g for 10 min and the supernatant was discarded. Finally, 2 ml of PBS was added to sperm pellet, vortexed gently for 3 s and was kept at 37° C into an incubator in humid condition containing 5 % CO2 in air for 30 min allowing sperm cells to swim-up into the medium. Before the experiment, sperm cells were aspirated carefully from upper layer of the stock medium and resuspended again into PBS to adjust the final concentration of 2.5 x 107 cell/ ml, evaluated by Neubauer counting chamber. For in-vitro experiments progressively motile spermatozoa >50% after dilution was considered to avoid non specific agglutination. Above procedures were performed at room temperature (35 ± 2° C) and completed within 2 h after slaughter. Preparation of test solutions To quantify the sperm agglutinating, immobilizing activities and titer of SF50, serial two fold dilutions of test solutions of different protein concentrations were prepared by dissolving SF50 with 0.15 PBS, PH 7.2 by double dilution (27) into different sterile screw-capped test tubes. Similarly, BSA solutions were prepared for the control study. Protein concentration for both the SF50 and BSA were adjusted to 10.95 µg, 5.47 µg, 2.73 µg, 1.36 µg, 0.68 µg, 0.34 µg, 0.17 µg, 0.08 µg, 0.04 µg and 0.02 µg respectively in per ml of PBS for the experiment and kept at 37° C prior to the experiment. Evaluation of spermatozoan agglutinability and motility The binding of SF50 to the sperm membrane thereby its ability to agglutinate spermatozoa were estimated semi quantitatively. For the evaluation, 20 µl each from prepared test solutions of different protein concentrations were taken into different Eppendorf tubes separately and 100 µl of sperm suspension was added (1:5) individually to each tube. Sperm-protein mixtures were vortexed gently. Observations were made under Leitz phase contrast microscope (x 200, x 400) immediately and after 20 s and 60 s respectively (28), placing 10 µl of each sperm-protein mixture onto microscopic glass slides. Two hundred sperm cells were counted randomly from 10 different microscopic fields to detect the number (%) of agglutinated spermatozoa as well as type of their agglutination. The sperm agglutinating activity of a sample was expressed as the highest fold of dilution that could agglutinate spermatozoa. Agglutination titer of SF50 on sperm cells was recorded on a quantitative scale from 0 (no agglutination) to 3+ (maximum to complete agglutination). The intensity (number) of sperm agglutination was calculated by haemocytometer as number of free spermatozoa before incubation (A0) and 20 s (A20) and 60 s (A60) respectively after the treatment. The percentage of agglutinated spermatozoa was calculated from 100 x (A0 – A20) / A0 and 100 x (A0 - A60) / A0. For photography, treated sperm suspensions were gently smeared on glass slides, dried up, fixed in formal-buffer solution and stained with PBS buffered solution of 3% Rose-Bengal. Spermatozoan motility was assessed microscopically (29). Two fold serial dilutions of SF50 solution, 20 μl each, was dispensed serially into different microtiter wells and fixed volume of sperm suspension, 100 μl, was added (1: 5) serially into each well and observed immediately under the phase contrast microscope (x 200, x 400) as above. A total of 100 spermatozoa were counted rapidly from ten different microscopic fields. The highest dilution of SF50 that displayed complete immobilization of sperm within 20 s was considered as maximum effective dose (28). The percentage of inhibition of spermatozoan motility was calculated by comparing with the BSA control. Spermatozoan motility recovery test Each treated sperm samples were washed and centrifuged twice with PBS at 500 g for 10 min and supernatant was discarded each time to remove the traces of SF50. To each sperm pellet buffered-glucose saline (0.2 g NaCl, 0.14 g NaHPO4, 0.14 g KH2PO4, 3 g D-glucose and double distilled water up to 100 ml, PH 7.4), 200μl, was mixed, vortexed gently and incubated in humid condition at 37 0C in a incubator containing 5% CO2 in air for 30 min. Spermatozoan motility was observed under Leitz phase contrast microscope (x 400) by placing 5 µl of each treated sperm suspension onto clean microscopic glass slides. Even if a single sperm within the 10 fields examined showed any sign of jerking or motility, it was considered as “positive”, and the values were expressed in percentage of motile and/or viable spermatozoa (30). All the experiments were repeated six times including controls and their mean results were considered. Statistical analysis All the data were analyzed by SPSS 10 (Statistical Package for Social Science) (31). Results Total protein content and pH value of SF50 were 109.5 µg/ ml and 7.4 respectively. SDS-PAGE of SF50 revealed that nearly 20 polypeptide bands had an estimated molecular weight between 90 kD to 16 kD in which 68, 64, 55, 53, 51, 46, 37, 34 and 23 kD proteins exhibited major bands and 90, 87, 84, 78, 72, 47, 45, 30.5, 28, 26, 28, 18 and 16 kD proteins had minor bands respectively (Figure 1). During observation, sperm samples were assessed for pattern and degree of SF50 induced agglutination as well as effect of incubation time on sperm agglutinability and motility of treated spermatozoa with different concentration of SF50 and BSA. Observations revealed nature of sperm agglutination were head-to-head type, i.e. agglutinated cells were adherent to each other mostly at the acrosomal region (Figure 2a and b). After supplementation of any concentration of SF50, sperm cells were started to agglutinate almost instantly. There was a constant tendency to enhance sperm agglutination in all the SF50 medium and the average percentage of head-to-head type agglutinated spermatozoa significantly increased (p<0.001) in a concentration and timed dependant manner. The extent of agglutination was related to protein concentration. In all the cases increased incubation period had more pronounced effect on the number of sperm cells that agglutinated. Higher number of head-to-head agglutinated spermatozoa (75.83 ± 0.55) were found after addition of 10.95 µg of SF50 within 20 s and increased (90.51 ± 0.4) over the incubation period at 60 s (Figure 2a), and lowest number of agglutinated spermatozoa 3.66 ± 0.45 and 12.58 ± 0.4 were found with 0.02 μg of SF50 at 20 s and 60 s (Figure 2b) respectively. All the values were highly significant (p<0.001) between the treatment and incubation period respectively (Table I). The aggregated sperm cells were not dissociated by agitating the suspensions. Observations also revealed that after supplementation of SF50, the pattern of intense forward motility of spermatozoa changed immediately to vigorous and whiplash like movement for a few seconds and sudden attachment of sperm heads occurred. With higher doses of SF50, spermatozoa were agglutinated and immobilized instantly. Maximum numbers of sperm cells were immobilized within 20s up to the use of 0.68µg SF50. However, with the other lower concentrations of SF50 sperm cells were also exhibited whiplash like movement instantly and after very few seconds sperm cells began to attach each other and clusters were formed, however, a good number of sperm cells exhibited either oscillated flageller movement, circular or irregular movement of the tail where sperm heads remained completely fixed and ultimately immobilized within 60s. No spermatozoan motility was recovered when SF50 treated samples were incubated in buffered-glucose saline. The BSA was observed to have no agglutinating or immobilizing effects on the sperm cells (Table I). Discussion The marine environment is exceptional reservoirs of bio-active natural compounds, many of which exhibit structural features that are not found in terrestrial natural compounds due to synthesize of unique secondary metabolites within the cells of marine organisms (32). Bioactive compounds from marine flora and fauna which contain various types of lectins are used as important tools for examining various modifications in the distributions of cellular glycoconjugates/cell surface receptors and the changes that occur in glycoconjugates during cell differentiation and maturation (33). Specific lectins bind to certain specific sugars. Due to sugar specificity the agglutination response of lectins is highly specific. As lectins possess high specificity to the accessible carbohydrate side chains, therefore, they are widely used as specific probes to investigate the cellular stages of differentiation, maturation, microenvironments and their architectures, and can reveal heterogeneity in the distribution of glycoconjugates. Moreover, lectin binding proteins exhibit changes during embryonic differentiation, cell maturation, aging, metaplastic alterations, malignant transformations, blood typing, mitogenic stimulation (34, 14). Present observation revealed that SF50 was able to agglutinate and immobilize the sperm cells which varied with the concentration of SF50 as well as influenced by the incubation period also. Probable reasons of these phenomena are not known clearly. However, Lectin induced agglutination is complex (34) and the molecular basis for the increased agglutinability is unknown. It could be that cell surface carbohydrates can act as cell-cell recognition molecules (35, 36) for agglutination of cells. Surface glycoconjugates on one cell may bind to complementary carbohydrate-binding protein (lectin) on another cells resulting in the initiation of a specific interaction. This may result in the adhesion of the two cells via carbohydrates and specific cell surface receptors. Hence, explanation of goat sperm agglutination might be formation of glycoconjugates with the sialic acid moieties contained in the membrane component of the spermatozoa (37) after addition of SF50 which is specific for sialic acid (23). Moreover, agglutination phenomenon induced by SF50 indicted perhaps SF50 possesses multiple sites or remain in the form of a multimer to bind sperm components resulting lectin-carbohydrate interaction. Furthermore, it was also presumed that SF50 plausibly capable to recognize specific carbohydrate domains present on the goat acrosomal membrane and are responsible for formation of glycoconjugate which could be an important factor for head-to-head type agglutination of sperm cells. Observation also revealed number of agglutinated spermatozoa increased over the time of incubation. Changes involved in SPM receptors induced by SF50 as well as SF50 binding site mobility of spermatozoa may occur during incubation. Increased agglutinability over the time may also happen due to capacitation of spermatozoa. Changes in lectin-binding or lectin induced agglutination associated with sperm capacitation have been described in model animal systems including mouse (38), hamster (39, 40), rat (41), rabbit (42, 43), guinea pig (6, 44-46) and ram (47). The cross-linked complexes formed between SF50 and its specific multivalent carbohydrates and glycoconjugates on the sperm surface were thought to be able to modify membrane permeability and to induce capacitation thus responsible for more agglutination. Observed sperm agglutination particularly in the acrosomal area also indicated that heterogeneous distribution of SF50-specific binding saccharide ligands may present only on the acrosomal surface. Heterogeneous distribution of sperm carbohydrate domains are also consistent with the finding of Nicolson et al (48) that sperm binding sites with Ferritin-lectin conjugates exhibited differences in the densities of lectin receptors in various sperm regions and changes in the same region occur during epididymal passage and after ejaculation in rabbit. However, it appears that SF50 receptors are present on the goat head plasma membrane. Therefore, SF50 induced sperm agglutination and increased agglutinability over the time might be due to changes and / or recognition involving receptors / carbohydrate domains of the sperm cell surfaces thereby formation of glycoconjugates induced by the SF50. Furthermore, it is important to determine whether the agglutination and immobilization of sperm cells are due to specific glycoproteins present on the SPM or of protein glycosylations which are present on the cell surface membrane components. The explanation for sperm immobilization induced by SF50 could be exocytosis of the SPM which require further study. The biochemical analysis of SF50 and SPM glycoproteins will help to understand the underlying mechanisms and importance of terminal sugar residues for the study of binding and immobilizing characters of sperm cells. Conclusion Present experiment reveals bio-active compound, SF50, isolated from marine cone snail Telescopium telescopium could be a novel biomarker to study the sperm surface and may prove an useful tool to differentiate in the structure compositions and for identification and characterization of mechanisms leading to male infertility. Moreover, sperm glycocalyx modification observed in this study could provide insights to the molecular modifications accompanying problems of subfertile goat and males of other species. More detailed pictures could be synthesized also on the sperm surface with the supplementation SF50 for different biological activities of male gametes during maturation, capacitation, acrosome reaction, fertilization, including sperm surface heterogeneity and for sperm integrity. Moreover, SF50 as an alternative source as local barrier contraceptive and/ or antifertility agent could be thought off. However, purification and characterization of SF50 and its application require more scientific approach. References
© Copyright 2010 - Iranian Journal of Reproductive Medicine The following images related to this document are available:Photo images[rm10002f1.jpg] [rm10002t1.jpg] [rm10002f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}