 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Iranian Journal of Reproductive Medicine Vol. 8, No. 1, Winter, 2010, pp. 45-50 The point mutations of m itochondrial tRNA threonine and proline in idiopathic repeated pregnancy loss Seyed Mohammad Seyedhassani1,2 M.D., Massoud Houshmand1 Ph.D., Seyed Mehdi Kalantar2 Ph.D., Abbas Aflatoonian2 M.D., Glayol Modabber1 M.Sc., Fatemeh Hadipour3 M.D., Mohammad Hossein Fallahzadeh2 Ph.D.
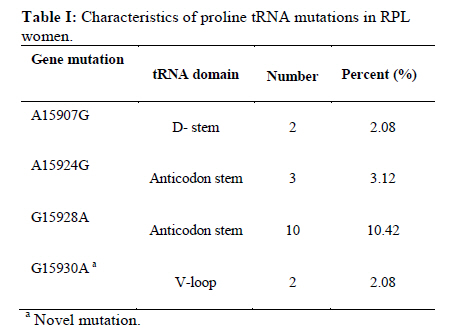
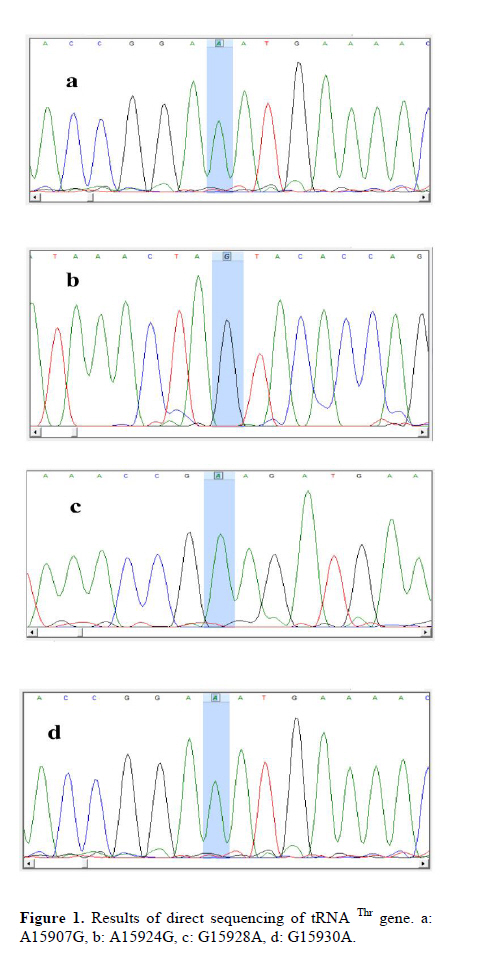
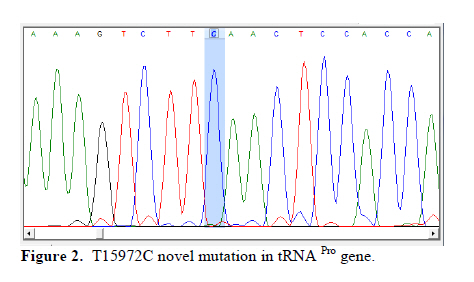
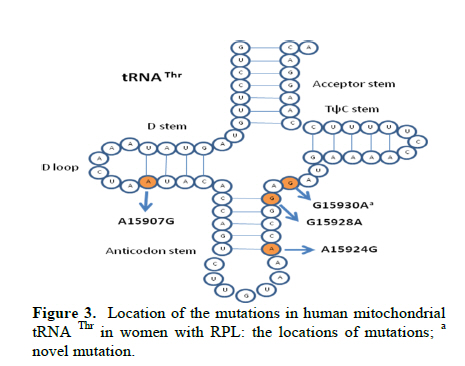
Corresponding Author: Massoud Houshmand, Medical Genetic Department, National Institute of Genetic Engineering and Biotechnology (NIGEB), Tehran, Iran. E-mail: massoudh@nigeb.ac.ir , housh62@yahoo.com Received: 30 January 2010; accepted: 14 March 2010 Abstract Background: Mitochondrial transfer RNAs (tRNA) genes are essential components of protein biosynthesis. These genes are hotspots for mutations. These mutations are associated with a wide spectrum of human disease. Many genetic factors are known in assessment of repeated pregnancy loss (RPL). Key words: tRNA, Mitochondrial mutation, Repeated pregnancy loss. Introduction Pregnancy loss is the most common complication of pregnancy and can be defined as the unplanned spontaneous loss of pregnancy before the fetus be able to survive extra uterine. Traditionally, repeated pregnancy loss (RPL) has been defined as at least three or more consecutive recognized pregnancy loss. Advances in the detection of early pregnancy revealed that about 70% of human conceptions fail to achieve viability (1); but clinically recognized pregnancy loss will occur in 15% of cases before 20 weeks of gestations (2). About 1 in 300 couples and 0.5-2% of women involved in RPL (3). Many etiological factors are known for RPL. Usually, most females with a chief complaint of RPL will be evaluated under the care of gynecologist for these possible causes. These factors are responsible for about 50% of RPL. They include uterine anomalies, chromosomal aberrations, infectious conditions and endocrine dysfunction such as luteal phase deficiency and hypothyroidism (4, 5). Genetic factors are actively being sought in idiopathic cases. Association studies have been done to help understand the role of candidate single genes toward the fetal loss. Until now, some polymorphisms have been suggested that increase the chance of pregnancy loss in women (6-14). A recent study revealed a higher frequency of mitochondrial DNA (mtDNA) variations in women with RPL (15). Mitochondrias are the bioenergetics and metabolic centers of the cells. During a process of high-energy consumption such as cell proliferation and development, the role of mitochondria and genome condition and competency are more important. This role is conducted through oxidative phosphorylation by producing ATP (16). Dysfunction in the mitochondrial respiratory chain causes a various group of progressive incurable diseases leading to severe disability and premature death (17). Increasingly, it is supposed that mitochondrial dysfunctions can cause oocyte wastage and early fetal loss by changing the activation of apoptotic process (18). The double strand circular mitochondrial DNA consists of 16,569 base pairs encoding 37 total genes in human: 22 tRNA , 2 rRNA , and 13 peptide genes (19). Since the first description of pathogenic mutations in the mitochondrial genome, over 200 disease-correlated point mutations and rearrangements have been found in association with a variety of mitochondrial cytopathies (19). More than half of these mutations have been located in tRNA genes that constitute 9% of the entire mitochondrial genome (20). Thus, mitochondrial tRNA genes are hotspots for mitochondrial pathogenesis and contribute in a disproportionate way to the etiology of disorders caused by mitochondrial DNA mutations, which is conceivable due to their central role in mitochondrial protein synthesis. Previously, the significant difference in the prevalence of spontaneous abortions was shown in the diabetic RPL patients with tRNA leucine mutation at position 3243 (21). In comparison, a little less than half of the mitochondrial mutations affect protein coding genes, which comprises 68% of the entire mitochondrial genome (22). We are describing mutations on tRNA threonine and proline in women with idiopathic RPL. Materials and methods This research was an analytical descriptive study. In total 96 women were been diagnosed as idiopathic RPL at a primary stage of evaluation. They were referred between September 2006 and June 2008 to the Research and Clinical Center for Infertility, Yazd University of Medical Sciences, Yazd, Iran. All of these women had past history of three or more consecutive pregnancy loss before 20 weeks of gestation. They had no history of live birth delivery. The known causes of RPL were evaluated. Diagnostic work up was consisting of uterine sonography, TORCH infections study (Toxoplasmosis, Rubella, Cytomegalovirus, Herpes Simplex virus type II and Listeria), and assessment of hormonal status, IgM and IgG anticardiolipin, antiphospholipids antibodies and, paternal and maternal karyotypes. The cases were explained and encouraged for consent and taking part in this study. The study was approved by ethic committee. A Flexigene blood DNA kit (DNA fast, QIAGEN, Cat. No. 51204) was used for isolating DNA from the blood samples based on the manufacturer’s protocol. The extracted DNA was kept at 4˚C. One primer pairs was used for amplifying the proline and threonine tRNAs coding regions. The 5’ end primer of this region (5’-ATC ATT GGA CAA GTA GCA TC- 3’) from nucleotide 15791 to 15810, and the 3’ end primer of it (5’-GCT CCG GCT CCA GCG TCT CG-3’) from nucleotide 91 to 110 was used to amplify this genome. The reaction mixture for PCR contained 7 pmol of each primer, 1unit of Taq polymerase (Cinnagene, Iran), each dNTP (Cinnagene, Iran) at a final concentration of 200 μM, and 2.5 μl PCR buffer at a final volume of 25 μl by distilled water. The reactions were done in thermal cycler (ASTEC- Japan). The PCR condition was as initial denaturation at 95˚C for 5min, then 35 cycles of denaturation at 94˚C for 60s, annealing at 58˚C for 60s, extension at 72˚C for 35s, and then final extension at 72˚C for 5min. The PCR products were evaluated on 2% agarose gel and then these fragments were purified and sequenced by Macrogen Company (Seoul, South Korea). The published revision of Cambridge reference sequence ( http://www.mitomap.org/ ) was used for comparing the results by the Chromas and Clustal X program. The sequence variants not found in the corresponding record of MITOMAP and other human databases were defined as novel variations. Results Our data showed that the Mean±SD age of the women with RPL was 28.73±5.86 years and the mean±SD for the gestational age at the time of miscarriages was 10.35±3.75 weeks. These women had the history of 3 to 11 miscarriages (median 3). PCR primers from both sides of the threonine and proline tRNA genome were used to amplify the related mitochondrial genome sequence. Direct automated sequencing of the PCR-amplified mtDNA was done. The sequence analysis of threonine tRNA revealed 4 mutations (Table I) (figure 1). Among them, G15930A mutation was novel at the V loop in 2 women. Substitution of G to A at nucleotide 15928 was more common and seen in 10 females (10.42%). The other two mutations were A15907G and A15924G that were found in 2 and 3 women respectively. One mutation was found in proline tRNA at nucleotide 15972 in one patient, which change T to C. This mutation was located in D-Loop Domain of this tRNA and has not been reported yet (Figure 2). Discussion The tRNAs are small ribonucleic acids (62 to 95 nucleotides) and essential components of protein synthesis because they function to transport amino acids to the ribosome, match them to the codons of mRNAs and facilitate their protein biosynthesis (23). In this study tRNA threonine and proline were investigated. In total, A15907G, A15924G, G15928A and G15930A substitutions among 65 nucleotides of threonine tRNA, but only one T15972C substitution in 67 nucleotides of proline tRNA were found. Mitochondrial tRNA Thr mutations The A15907G mutation is located at the D-stem domain of tRNA Thr (Figure 3). As our knowledge this mutation is not reported in RPL patients or in human-related diseases. In one study, this mutation was reported in 2% of a normal control group as a variation (24). The A15924G in anticodon stem of tRNA Thr was seen in 3 women with RPL. This mutation is reported in some diseases such as: mitochondrial encephalopathy (25), Parkinson’s disease (26), Idiopathic cardiomyipathy (27) and fatal infantile respiratory enzyme deficiency (28). The other mutation in anticodon stem was G15928A that was seen in 10 women and was the most common mutation in tRNA Thr in our study. Also, the recent mutation is reported in Parkinson’s disease (26) and multiple sclerosis patients with severe optic involvement (29). The nucleotides in anticodon are the least affected bases because it has been identified only one mutation in one tRNA affects any of the three bases necessary for decoding(30). Substitutions at these locations could cause cellular outcomes too severe to sustain cell growth. The G15930A in V-loop or accessory stem was seen in 2 cases. Although it seems the effect of this substitution is less than the others, however, identifying its role needs more evaluation. The other mutations which are reported previously consisting of: T15908C in deafness-associated 12S rRNA A1555G mutation (31), G15915A in mitochondrial encephalomyopathy (32, 33), A15923G in lethal infantile mitochondrial myopathy (25), fatal infantile respiratory enzyme deficiency (28), newborn cardiopulmonary arrest (34), G15927A in Parkinson’s disease (26), multiple sclerosis patients with severe optic involvement (29), deafness-associated 12S rRNA A1555G mutation (35), G15950A in Parkinson’s disease (36) and A15951G in LHON-associated ND4 G11778A mutation (37). Mitochondrial tRNA Pro mutation The T15972C mutation is found at the D-loop domain of tRNA Pro. There are several reports of this area as A15965G mutation in Parkinson’s disease (37), C15975T in late-onset ataxia, retinitis pigmentosa, deafness, leukoencephalopathy and complex I deficiency (39), C15990T in myopathies (40), G15995A in cystic fibrosis (41) and T16002C as a novel mutation (42). However, T15972C is seen in 1 woman with RPL and is not reported yet. Two pathways for inducing the mtDNA including tRNA mutations are considerable. It is demonstrated that the disorder of mtDNA can be induced by the defects of nuclear DNA (43). Another promising pathway is associated with the reactive oxygen species (ROS). The mitochondrial genome is extremely susceptible to damages from continuous exposure to ROS. It has been suggested that the ROS is produced endogenously from mitochondrial respiratory chain and have been considered to be involved in the increased ratio of point mutant mtDNA (44). It is thought that the condition is probably induced by the inhibition of the repair system for ROS mediated damage to mtDNA, detoxification of ROS, or increase in ROS production and might be possible causes for tRNA point mutations of mtDNA. The various aspects of tRNAs function and the effect of their different mutations have been evaluated. Disease-related point mutations could potentially influence mitochondrial tRNA and affect their primary, secondary, and tertiary structure. It leads to protein synthesis defects and, in turn, mitochondrial dysfunction. Ultimately, these disturbances result in cellular dysfunction which is more important in cell proliferation and development. Embryo as a main material in protein synthesis and related functions can be more sensitive to these alterations leading to wastage. Conclusion Further study of an expanded series of these tRNA mutants is recommended in order to create a consensus or framework. It would permit the description of the cellular and phenotypic effects of tRNA mutations in related diseases. Because of difficulties to get aborted materials, it was focused on parents. More studies are necessary to clarify and show the primary or secondary role of tRNA mutations in embryonic development. Acknowledgment The authors would like to thanks IVF teams of Yazd Research and Clinical Centre for Infertility, the laboratory team of National Institute of Genetic Engineering and Biotechnology and also Yazd clinic of genetics for their collaboration in this study. References
© Copyright 2010 - Iranian Journal of Reproductive Medicine The following images related to this document are available:Photo images[rm10008t1.jpg] [rm10008f1a-d.jpg] [rm10008f2.jpg] [rm10008f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}