 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Iranian Journal of Reproductive Medicine Vol. 8, No. 3, Summer, 2010, pp. 101-110 Effect of bilateral uterine artery ligation on follicular atresia in ovaries of mature female rabbits; histomorphometric and histochemical study Mazdak Razi1 Ph.D., Kaveh Akhtari2 D.V.M., Ali Reza Najafpour2 D.V.Sc., Keyvan Abdi2 D.V.Sc., Rasoul Shahrooz1 D.V.Sc., Simineh Shahmohamadloo1 D.V.M., Sajad Feyzi1 Ph.D., Hadi Cheragi1 D.V.M.
Corresponding Author: Mazdak Razi, Department of Histology and Embryology, Faculty of Veterinary Medicine, Urmia University, Urmia, Iran. Email: mazdak_razi22@yahoo.com Received: 4 January 2010; accepted: 14 April 2010 Code Number: rm10019 Abstract Background: Nowadays it is proofed that the uterine artery plays essential role in follicular growth and/or post parturition hemorrhagic. Key words: Atresia, Carbohydrate, Lipase enzyme, Lipid foci, Ovary, Rabbit, Uterine artery. Introduction In rabbits, the ovaries possess a dual blood supply, including ovarian and uterine arteries. The ovarian bifurcation of the uterine artery creates utero- branch, which supplies the tip of the uterine horns, the oviduct and also forms anastomosis with a primary branch of the ovarian artery. Thus there is a links between the uterus and the ovary by the vascular junction (1). Previous studies showed that the uterine artery is the major source of the ovarian blood flow in cycling Rhesus monkeys (2) and humankind (1-5). It is very important to understand the contribution this anastomosis makes in supplying blood to the ovaries. Any disturbance that influence this correlation, leads an adverse effect on the ovarian follicular growth and hormonal balance (3). Different methods such as, total hysterectomy, unilateral hysterectomy, tubal ligation, and total salipingectomy can destroy different parts of the tube and block the correlation between the uteri and the ovary (3-6), leading to reduce the ovarian blood supply, thus it can cause decreasing of ovulation rate (7,8). For example, selective uterine or hypogastric (internal iliac) artery ligation and mass uterine artery ligation are surgical options for avoiding hysterectomy in intractable postpartum hemorrhage (9), while this option can lead to shorten corpus luteum life span and also cause significant changes in the rate of hormones in animals (4,10) and human (5,7). On the other hand, technological advances and increasing social acceptance has made tubal sterilization the most commonly chosen form of contraception for women, especially during the last decade of their reproductive life (2, 12, 13). The possible adverse influence of tubal sterilization on ovarian function has been understood during recent years (2, 13). Studies investigating the probable cause of the ovarian disturbance have especially focused on the ovarian arterial blood supply at the main arterial level (such as the ovarian artery). However the ovarian stroma obtains its blood from anastomosis as well as ovarian artery. Previous studies could not completely define the exact relationship between the follicular atresia (depending on follicular size) and surgical intervention to the ovarian blood supply. Thus the first purpose of the current study was to investigate the follicular fate in different sizes. According to this fact that during the first stages of the apoptosis, pathological atretic follicles show different patterns for cytoplasmic carbohydrate and lipid ratio, the second purpose for the present study was to determine the probable histochemical changes which occur in the rate of the carbohydrate and lipids in the ovaries of the experimental rabbits as a laboratory model and to compare these features with the control-sham cases. Materials and methods
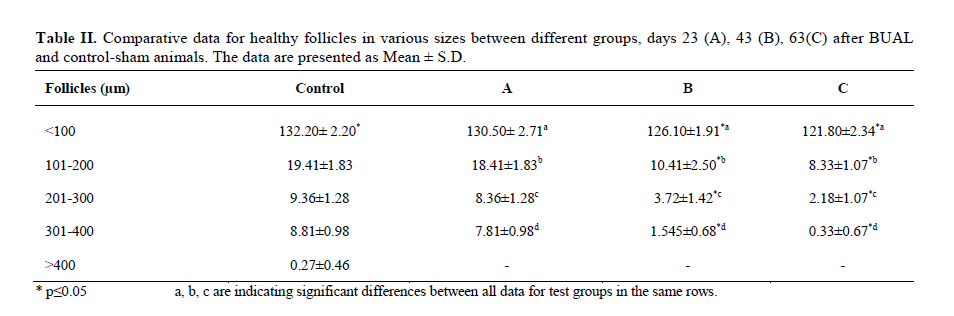
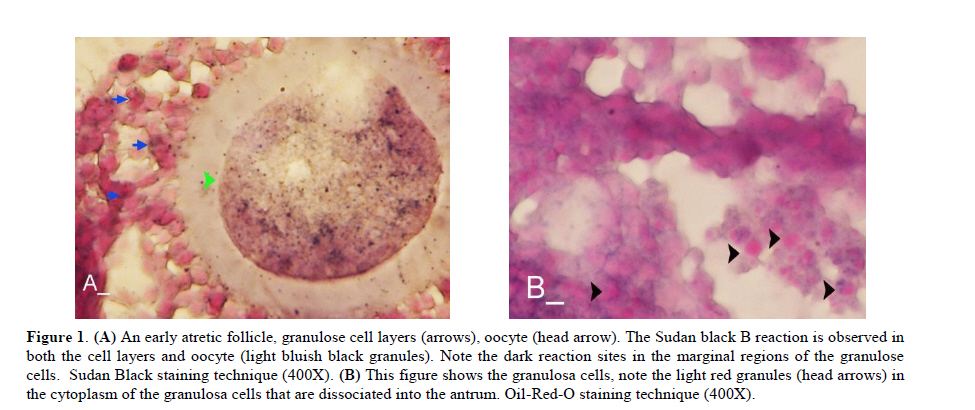
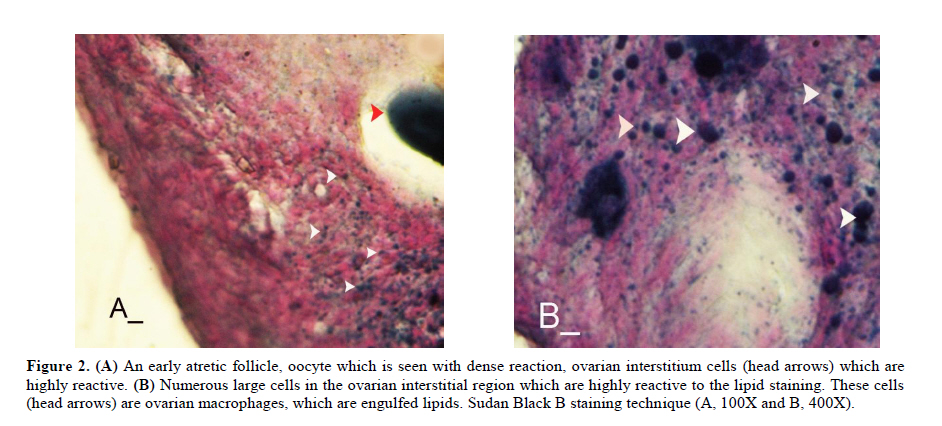
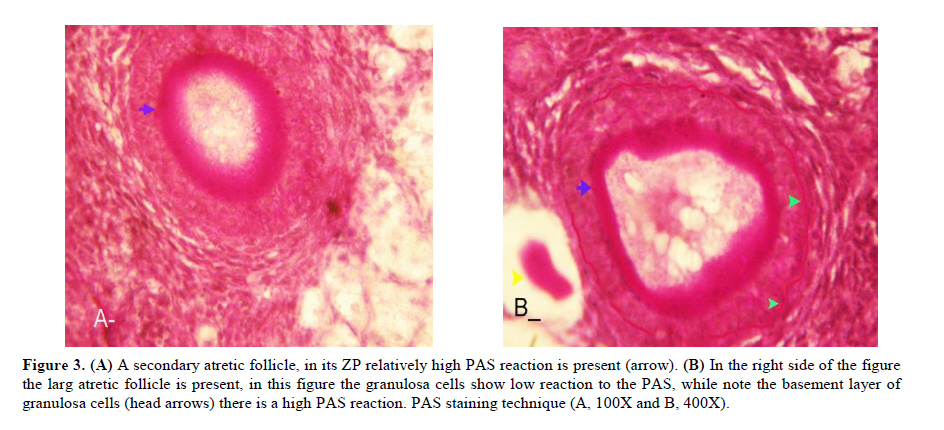
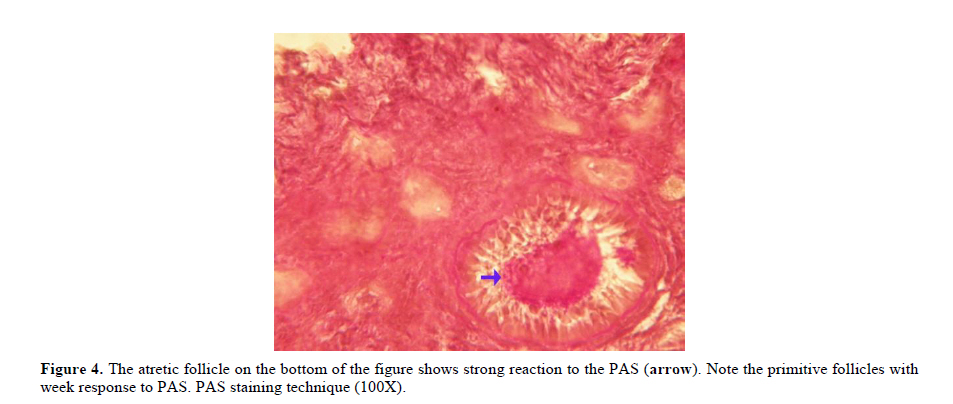
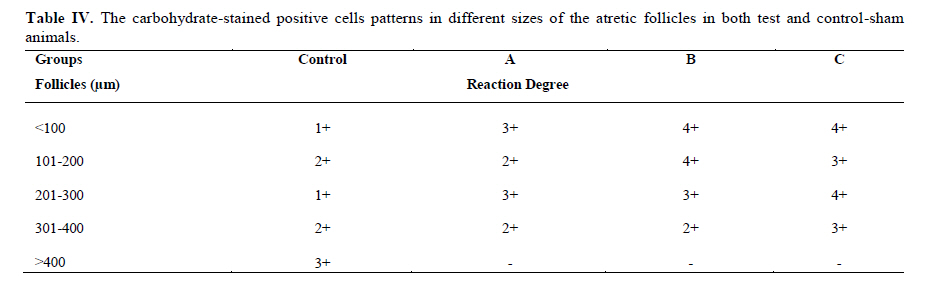
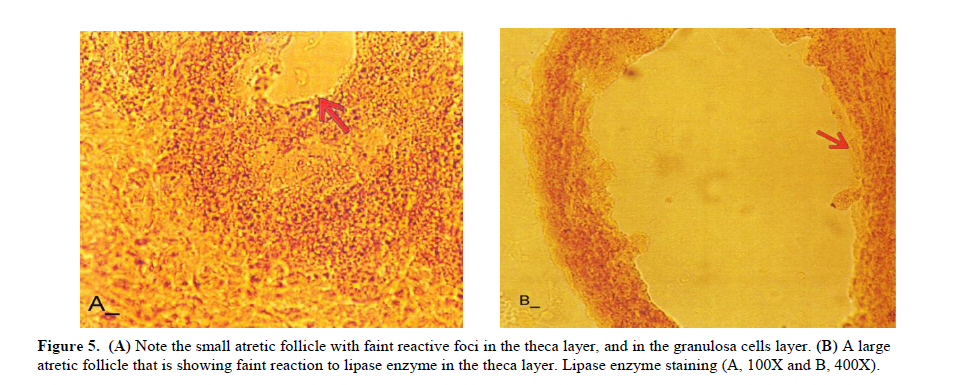
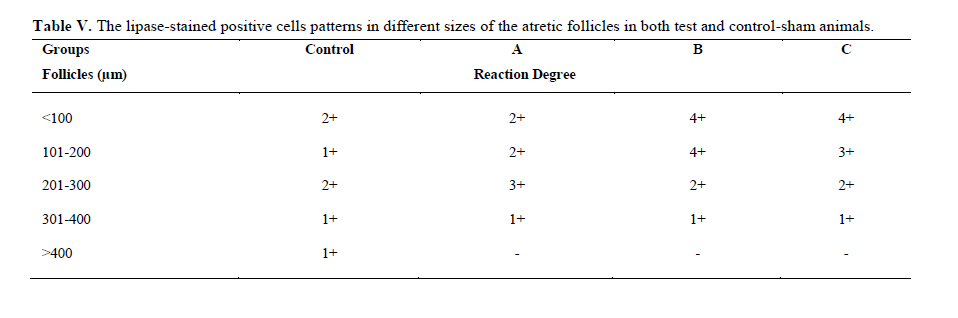
Twenty four rabbits (Albino-albino), 5-6 months old were selected in order to perform an original study on female rabbits. The rabbits were obtained from the animal centre of Faculty of Veterinary Medicine, Urmia, Iran. The animals were acclimatized in an environmentally controlled room (temperature 17- 23°C; relative humidity 50-70%; light on 12h/12h dark). Food and water were given at libitum. The rabbits were assigned into two groups of test and control-sham groups. Test group was subdivided into 3 groups (6 rabbits for each group), on days 23 (Group A), day 43 (Group B) and day 63 (Group C), after the uterine artery ligation animals were ovariohysterectomized. The vulva of these animals were observed daily for color intensity, based on the rapid and consistent reddish/purple, to determine day 0 of the surgery in test groups (4). Approval for this study was gained from Urmia University of Veterinary Medicine Animal Care and Ethics Committee. Surgical method for bilateral uterine artery ligation Test and control-sham groups were anesthesiated by Ketamin 5% (Germany/Trittau) 35 mg/kg, and Xylazine 2% (Holland/Woerden) 5mg/kg intra-peritoneally. After anesthesia, the rabbits were positioned in dorsal recumbency, with hind limbs restrained in extension. An incision (2-3 cm) in midline was made through the skin, between the ambilicus and the cranial rim of the pelvis (Pelvix symphisis). The isolated opposite structures were carefully inspected. The uterine arteries bilaterally ligated (by 0-2 silk) close to the splitting region of the uterine horns. No arterial ligation was conducted in the control-sham rabbits. Ovariohysterectomy On days 23, 43 and 63 after the ligation, ovariohystrectomy was performed in the test groups of A, B and C, respectively. Animals were anesthetized as previously described. After incision, the body of the uterus was isolated and clamps were placed. The ovarian pedicles were isolated and ligated and the uterine artery and vessels supplying the broad ligament were also ligated. Finally, organs dissected out and the remaining stamps were checked for hemorrhages, before placing them back through the incision. Histomorphological analyses On days 23, 43 and 63 after operation, the ovaries were removed and fixed in formaldehyde acetic solution (IFAA, Merck, Germany) for 4 weeks. Ultimately, they were dissected free from per-ovarian tissues. One half of the samples were processed through paraffin embedding and cut with rotary microtome (Microm GmbH, Germany) serially and stained with Hematoxyline and Eosin (Merck, Germany) and PAS (Pajohesh Asia, Iran) techniques. Histomorphometrical study was performed by measurements of follicles that they were characterized in terms of their sizes to under 100, 101-200, 201-300 and 301-400 and larger than 400µm. Study of lipids For the study of lipids, we adapted Oil-Red-O (Pajohesh Asia, Iran) and Sudan Black B (Pajohesh Asia, Iran) techniques. In this procedure, the tissue specimens were fixed in 10% neutral formaldehyde solution and sectioned with cryostat microtome (Bright 00361, England) and stained through special techniques for each of them separately. The atretic follicles with 20% positive lipid-stained cells were considered as pattern one (1+), with 50% positive lipid-stained cells were considered as pattern two (2+), with 60% positive lipid-stained cells were considered as pattern three (3+) and with 100% positive lipid-stained cells were considered as pattern four (4+). Study of Carbohydrates For the PAS, we adopted paraffin sectioning procedure and stained through special technique for the study of carbohydrates. The atretic follicles with 20% positive carbohydrate-stained cells were considered as pattern one (1+), with 50% positive carbohydrate-stained cells were considered as pattern two (2+), with 60% positive carbohydrate-stained cells were considered as pattern three (3+) and with 100 % positive carbohydrate-stained cells were considered as pattern four (4+) (14). Lipase enzyme For the investigation of lipase enzyme in the ovarian tissue, i.e., ovarian follicles and ovarian interstitial tissues, the fresh unfixed specimens were sectioned with cryostat microtome (frozen sections), and stained through special techniques for the detection of this enzyme. The atretic follicles with 20% positive lipase-stained cells were considered as pattern one (1+), with 50% positive lipase-stained cells were considered as pattern two (2+), with 60% positive lipase-stained cells were considered as pattern three (3+) and with 100% positive lipase-stained cells were considered as pattern four (4+). Statistical analysis All results are presented as mean ± SD. Differences between quantitative histological data on days 23, 43 and 63 of the treatment were analyzed with two-way ANOVA, followed by Bonferroni test 4.00. p<0.05 was considered as significant differences. Results BUAL increases the rate of the atresia on the ovaries The ovaries from control-sham group showed many follicles in various stages of development including primary, secondary and tertiary follicles with different sizes from <100 µm to > 400 µm, whereas there were no large antral follicle (>400 µm) in each histological section of the test groups. BUAL resulted in a significant (p<0.05) decline in follicular size in test groups in comparison to the control-sham group. The cortex of the ovaries covered with small antral atretic follicles (largely <100-200µm) in the test group. This situation advanced depending on the time in BUAL cases. Accordingly, atresia was manifested mostly in <100 and 201-300µm follicles in the test groups. These follicles were atresiated remarkably on day 63 in comparison to other days. In test groups, total atretic follicles were higher than control- sham. On day 23 after operation, follicles <100 µm and on days 43 and 63, 101-200 µm follicles showed the highest rate of atresia indicating 39.04%, 40.90% and 38.46% respectively in comparison to the other sizes. The data for atretic follicles are presented in table I. Our observation showed that there were follicles larger than 400 µm in control-sham animals while no such large follicles were observed in the test groups. In comparison of normal follicles, the observation revealed a significant decrease in normal follicles in different sizes in test groups. This situation was progressed time-dependently, which on day 63 ovaries showed the lowest normal follicle, especially in size between 301-400 µm follicles. The highest rate of normal follicles was observed in group A of test groups. The data for normal follicles are depicted in table II. BUAL induce lipid foci in the ovaries Histochemical staining for the lipids revealed the lipid foci in the cytoplasm of some cells such as follicular granulosa cells, follicular theca cells, oocyte and macrophages in the form of cytoplasmic droplets on the ovaries in the operated (BUAL) rabbits (Figure 1 A/B). In the granulose cells of the un-advanced atretic follicles, the Oil-Red-O and sudan-black reaction was stronger in comparison to the advanced atretic follicles. Some of the interstitial cells of the BUAL induced rabbits showed considerable response to the Oil-Red-O and sudan black staining techniques (Figure 2 A/B). It should be considered that these cells were observed in the form of large cell colonies that were stained dense bluish-black in the sudan black B staining technique. The distribution of these cells was higher in the BUAL rabbits in comparison to the control-sham group. This situation was advanced by the time in the test group. Accordingly, histochemical analyses of the ovaries of group C, showed the highest presentation of the Oil-Red-O and sudan black positive reaction sites in the form of cytoplasmic droplets in the interstitial cells. In contrast to the atretic follicles, the normal follicles showed negative Oil-Red-O and sudan black reaction. On the advanced stage of the atresia the histochemical staining for saturated lipids revealed remarkable reaction in the follicular granulosa cells in comparison to the theca cells. The oocytes showed stronger reaction to the lipids histochemical staining determination technique in the first stages of the atresia in comparison to the advanced stages. The data for lipid foci accumulation are presented in table III. BUAL causes the positive PAS reaction on the ovaries This reaction was positive in structure of atretic follicles such as ZP, basement membrane of granulose cells, stroma of follicles, follicular fluid and granulosa cells (Figure 3 A/B). In contrast, the normal follicles in the sham-control rabbits showed very faint response to the PAS. The small atretic follicles which were committed to the precautions atresia showed remarkable PAS reaction in the antrum. The ovarian macrophages were manifested with remarkable PAS reaction sites in the cytoplasm. The granulosa cells were observed with PAS positive intra-cytoplasmic reaction sites in the precautious atretic follicles. PAS reaction was positive in the interstitial tissue between the granulosa cells of the atretic follicles. In some of the small atretic follicles, the oocyte showed intense PAS reaction that was advanced to more follicles by the time on the BUAL rabbits ovaries (Figure 4). The data for PAS-stained positive cells are presented in table IV. The PAS reaction was also seen in ovarian stromal connective tissue, intima of blood vessels and ovarian macrophages. In the process of follicular atresia and follicular cyst formation, by the enlargement of such follicles, the PAS reaction was getting weaker progressively. BUAL elevates the reaction sites for lipase enzyme staining in the early stages of the atresia The atretic follicles, granulosa and theca cells were reactive for this enzyme. However, by progression of follicle atresia formation, this reaction became weak. The interstitial tissue cell showed faint lipase reaction. The lipase reaction in the theca layers of cystic follicles was quite stronger than the granulosa cell layers of these follicles (Figure 5). The data for positive lipase enzyme-stained cells are showed in table V. Discussion The purpose of the current study was to show the histological and histochemical features of the atretic follicles which are produced due to BUAL. We showed a significant enhancement of the atretic follicles formation which was accompanied with remarkable decrease in the rate of normal follicles. Previous studies showed that BUAL decrease the ovulation in rats (15, 16). During the ovulation, distinct homodynamic and structural changes occur in the ovary. An elevation in intrafollicular blood flow (17), increased capillary permeability (18), and breakdown of the extracellular matrix at the apex of the follicle (19) are major components of the ovulatory process. The ovarian blood flow is comparatively high and increases several folds during the ovulation (20, 21). Recent studies indicated a rapid and persistent reduction of blood flow after both the ovarian and uterine artery ligation, independent to the stage of the ovulatory process (10). According to Ahn et al, premenopausal total abdominal hysterectomy accelerated the ovarian dysfunction and the women treated with mentioned method are at the risk of early menopause and should receive adequate hormone replacement therapy (7). In another study it was reported that 6 months after hysterectomy the cortex of the ovary completely covered by corpora lutea, and there were many cystic and atretic follicles with few normal follicles presentation. Our results confirmed and extended the previous studies that on day 63 after BUAL the cortex of the ovaries were covered mainly with atretic follicles in different sizes. Moreover corroborating of previous studies in the present study histological observations revealed that the rate of ovulation decreased in the cases that under went to BUAL, because the rate of early atresia increased and this condition substantiated by the time. According to increased atresia in the BUAL cases and also absence of > 400 µm follicles in this study, it seems that BUAL causes significant hypo- ovulation. According to the previous reports (22), histochemical alterations are taken place in the extracellular matrix of the ovaries and in the follicular wall as well. Lobo and co-workers reported that the alterations in the amount of the collagens of the theca internal and external, the acidic glycosaminoglycans of the cystic and atresiated ovaries and carbohydrate alteration in granulose cells in the atretic follicles are more considerable in comparison to the normal follicles (23). Thus, histochemical study of the ovaries that under went to a remarkable atresia following BUAL may help to come close to answer some questions in this regard. Our results revealed that PAS reactions were present in atretic follicular structure such as ZP of the oocyte, basement membrane of the granulose cells, stromal tissue, follicular fluid and granulose cells (Figure 3). Microscopical analyses of different sizes of the follicles showed higher PAS reaction in granulosa cells and basement membrane of atretic follicles. This situation suggests that accumulation of glycoprotein or carbohydrates contents are present in such follicles. In the current study, the most considerable point was that in the process of the atresia the PAS reaction gradually became faint. In other study that was carried out on the large atretic follicles, the results indicated that the granulose cells showed some structural changes such as vacuolation of the marginal region of the cytoplasm, cytoplasmic lipid accumulations and piknotic nucleus (24, 25). In corroborating of previous studies, in the present research, the Oil-Red-O and sudan black B staining techniques showed heterochromatic nucleus with positive lipid reaction foci in the cytoplasm of the granulose cells in the atretic follicles. In the other study that was conducted on the estradiol-valerate induced polycystic ovary rats, the Oil-Red-O and sudan black B reactions were weaker in advanced cystic and atretic follicles in comparison to the early stage of the atresia (26). In the apoptotic process, an accumulation of the lipid droplets takes place in the cytoplasm of the follicular cells and this situation does not process in the advance stages of the atresia. Some of the authors believe that because of massive cell degeneration and removal, this reaction does not take place (24- 27). In the accommodate of the previous studies, our results revealed that in the first stages of the precocious atresia in the BUAL group, some structures of the atretic follicles had high reaction to the sudan black B and Oil-red-O staining technique. Recently, Blasco et al, (2004) reported that macrophages have critical role in regulation of the ovarian function. Macrophages have an essential role in phagocytosis of cellular residuals and participation in defense mechanisms in different organs (19). Previous studies reported that the ovarian macrophages have different origin. They reported that macrophages in the ovaries originate from granulose cells (27, 26). In the present study, macrophages were manifested in different regions of the ovaries in test groups. No macrophages were observed close or around the normal follicles, while they were localized with high density around the atretic follicles. It seems that the granulose cells dissociation and floating in the antrum leads to generate macrophages with granulose cells origin. In the present study, interestingly the presence of the ovarian macrophages, revealed by Oil-Red-O and sudan black B method, was higher in test group compared to the control- sham groups. More investigations showed that this situation advanced by the time, accordingly the group C rabbits revealed higher population of the macrophages with dense stained lipid droplets in the cytoplasm. It can be concluded that in response to high cell destruction in the atretic follicles and for the removal of these cells, the population and activity of the macrophages were increased accordingly. According to the earliest studies, the germinal epithelium of the rat ovaries always contained no lipase. In the primary follicles, neither the ovaries nor the follicular cells contained any lipase reaction. In the secondary and Graafian follicles, the ovaries which differed extremely in lipase activity were found. Follicular cells showed no lipase reaction excepting that the cells of the cumulus oophorus in the large Graafian follicles frequently showed a weak reaction of it (28). In the other study, the activity of the lipase enzyme was detected in the granulose and theca cells of the cystic and atretic follicles in the early stages of the atresia (26). Regarding the previous reports, in the current research, in advanced stages of the atresia, the lipase activity was very weak. While in the first stages of the atresia in the early atretic follicles, we saw high lipase activity in the cytoplasm of the granulosa cells. Accordingly, in the group C of the present study, observations demonstrated low activity of the lipase enzyme in granulosa and theca cells. It can be concluded that the biological activities of follicular cells in the process of the atresia were decreased and accordingly the normal life of them was also affected. Conclusion According to increased atresia in different sizes of the follicles (pre- antral and/or antral) in the case of BUAL and also absence of > 400 µm follicles in this study, it seems that BUAL caused significant hypo-ovulation by increasing the atresia. Referring to our results, accumulation of glycoproteins or the carbohydrates contents in the follicular cells and basement membrane of the atretic follicles was remarkable but the PAS reaction became faint by the time that the atresia was progressed. The apoptotic process can lead to the cell death because of the biological activities missing. In order to obvious increasing of lipid droplets in the follicular cells in the first stages of the apoptotic process (that occurred in the process of the atresia) probably the activity of the lipid enzyme increased accordingly, while by the advancement of the atresia and decreasing of the biological and biochemical activities of the follicular cells and altimetry destruction of these cells, the lipase enzyme activity decreased by the time. Acknowledgment The authors wish to thanks the staff of Histology and Embryology Laboratory, Mr. Ali Karimi for his helps, in processing of this study. This study was supported with Research Central of the Histology and Embryology of Urmia Faculty of Veterinary Medicine. References
© Copyright 2010 - Iranian Journal of Reproductive Medicine The following images related to this document are available:Photo images[rm10019f4.jpg] [rm10019t4.jpg] [rm10019f5a-b.jpg] [rm10019f1a-b.jpg] [rm10019t1.jpg] [rm10019t3.jpg] [rm10019t2.jpg] [rm10019f3a-b.jpg] [rm10019t5.jpg] [rm10019f2a-b.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}