 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
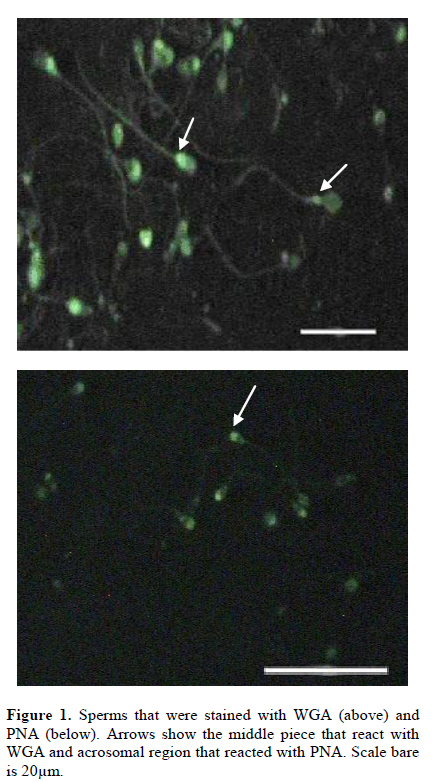
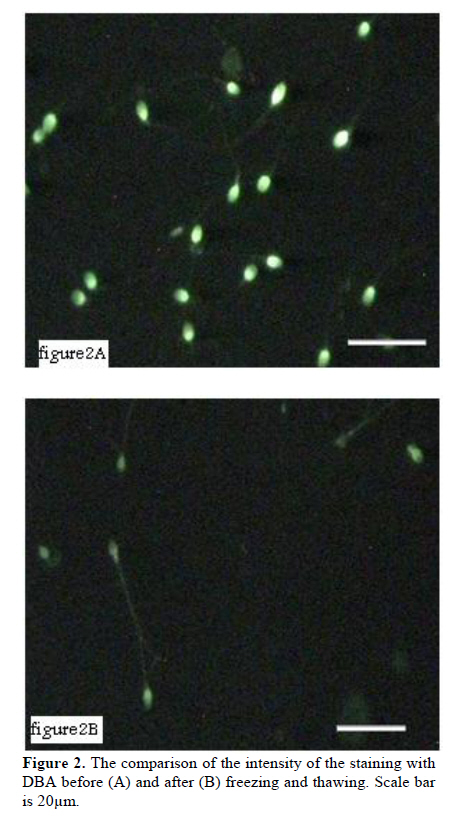
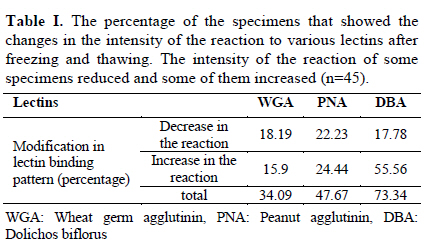
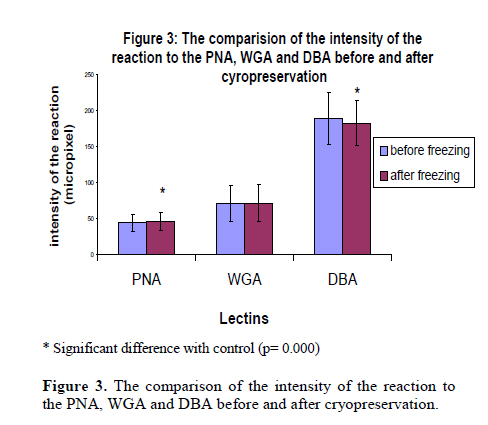
Iranian Journal of Reproductive Medicine Vol. 8, No. 3, Summer, 2010, pp. 119-124 Effects of cryopreservation on plasma membrane glycoconjugates of human spermatozoa Tahereh Talaei Ph.D., Tahereh Esmaeelpour Ph.D., Fatemeh Aekiyash B.Sc., Soghra Bahmanpour Ph.D. Department of Anatomy, Shiraz University of Medical Sciences, Shiraz, Iran. Corresponding Author: Tahereh Talaei, Department of Anatomy, Shiraz University of Medical Sciences, Shiraz, Iran. Email: talaeit@sums.ac.ir Received: 31 August 2009; accepted: 14 April 2010 Code Number: rm10021 Abstract Background: Cryopreservation has some detrimental impacts on sperms surface molecules. Modification of the sperm surface molecules can affect on fertility rate. One of the important surface molecules are glycoconjugates. Key words: Sperm, Cryopreservation, Surface Glycoconjugates, Lectin. Introduction The surface of all eukaryotic cells consists of a glycocalyx that is a carbohydrate-rich zone. This cell coat contains oligosaccharide chains that covalently bound to integral protein of plasma membrane (glycoproteins) or lipids (glycolipids), as well as polysaccharide chains that covalently linked to a protein core (proteoglycan). Like the glycocalyx of somatic cells, the glycocalyx of spermatozoa is critical for many specific cell functions (1). The sperm glycocalyx represents the primary interface between the male gamete and its environment. It is known that the glycocalyx involves in immunoprotection in the female genital tract, acquisition of fertilizing ability, acrosome reaction, and early gamete interactions (2). The presence of sialic acid and Galactose/N acetylgalactose amine (GAL/GAL Nac) containing glycoconjugate sequences were detected on the plasma membrane of the motile human spermatozoon (3). It has been shown that WGA (wheat germ agglutinin) could bind to almost all motile spermatozoa (3). Acrosome also contains a matrix that is rich in carbohydrates (4). The post-translational modifications in the glycosylation status have occurred during epididymal passage and this modification is significant in fertility. More N-Ac-galactosamine, N-Ac-glucosaminylated, and fucosylated residues were found on sperm membranes when they passed through the proximal part of the epididymis (5). These modifications may show the importance of the surface glycoconjugate in sperm normal function. There is evidence revealed that the modification of glycocalyx of sperm is related with male infertility (6). Mammalian spermatozoa were among the first cells to be successfully cryopreserved (7). However, both freezing and thawing expose the cells to severe stresses (8). It has been reported that cryopreservation impairs sperm motility (9, 10), viability (11), penetration into cervical mucus, penetration of zona-free hamster eggs (9), acrosome structure (10), and activity of acrosome protease and acrosin (12). During the cryopreservation process, osmotic effects of cryopreservation damage sperm cell membrane and reduce the fertilizing capacity (13). Cryopreservation may modify the sperm glycocalyx by physical damage. Therefore, cryopreservation may change the charbohydrate composition of the sperm membrane which is important for physiological functions such as capacitation (14), sperm membrane permeabilization, acrosomal reaction (3) and fertilization (15). There is evidence that revealed the modification of glycocalyx of sperm was related with male infertility (6). At least to some extent, the reduction in fertility capacity of cryopreserved sperms may be attributable to glycoconjugates modification. Because, the structural and functional integrity of the sperm plasma membranes are critical to fertilization, this study was design to determine if the content of the glycocalyx changes, after standard freeze and thaw procedure. Materials and methods Semen samples were collected from 45 healthy male, attending the cytogenetic clinic in Shiraz, Iran. All samples were assessed as normal according to WHO guidelines (16). The sperm count, motility, morphology, viability, the number of WBC, pH and also liquefaction state of semen were normal (16). Semen samples were allowed to liquefy at room temperature for 30 minutes and washed with medium. Sperm smears were prepared from each sample in the ply L lysine (Sigma, Cat No P4832) treated slides twice. The first smear from each sample was prepared before freezing (as control) and the second one was done after thawing. Freezing and thawing: The freezing of the sperm samples were done by cryopreserved media (Lifeglobal, Cat No LGSF-0707) according to manufacture instruction. Briefly, one milliliter of each sample was mixed with 0.7 ml of cryoprotective media (Human sperm preservation medium contains 15% glycerol in aqueous solution containing sodium choloride, sodium phosphate, calcium lactate, Human serum albumin, potassium chloride, sodium bicarbonate, glycine, sucrose, magnesium sulfate, glucose and HEPES) while swirling. The mixtures were allowed to equilibrate at room temperature for 10 minutes. The samples were drawn into 0.25 ml freezing straws with 1.5 cm of air at each end of them. The straw was later sealed and frozen in the liquid nitrogen vapor. The straw was placed in the horizontal position at 5 cm above to the surface of liquid nitrogen for 15 minutes. Then, it was quickly plunged into liquid nitrogen for storage. After 48 hours of cryostorage, the straws were taken out and thawed at 37°C for 10 minutes. Sperms smears were prepared from each sample in a ply L lysine treated slides. Lectin histochemistery: Before staining the smears with lectins, they leave to equilibrate at room temperature for 1 hour. The samples were washes in PBS that contained CaCl2, MgCl2 and MnCl2 (Merk). The samples were incubated in FITC conjugated lectins (10µm/ml) for 2 hours and washes for 30 minutes. The samples were mounted with antifade and observed by florescence microscope using a 490-nm excitation filter and a 530 nm barrier filter on the same day (5, 17, 18). The sperms smears were stained with lectins WGA, PNA and DBA (Sigma, Cat No L4895, L9142 and L7381) that labeled sialic acid, Galactose/N-acetylgalactoseamine and N-acetyl-D-galactosamine terminal sugar, respectively. WGA was also reacted to glucoseamine. The smears were fixed with buffer formalin, and were frozen until used. Photographs were taken from ten fields of each sample and the intensity of the reaction of each sperm in the fields was evaluated by Jama image analysis software. The brightness of the sperms heads were recorded by micropixel and was a number between zero and 255. The means of the brightness of sperms head after thawing was calculated and compared with those before freezing. The percents of the specimens that revealed modification in the intensity of the reaction were calculated. Staining with acridine orange: Some smears of each sample were fixed in Carnoy’s solution (methanol:acetic acid = 3:1) overnight. The acridine orange (AO) (Sigma Cat No 01662) staining solution was prepared as follows: 1% AO in distilled water was prepared and 10 ml of the solution was added to a buffer that was composed of 40 ml of 0.1M citric acid and 2.5 ml of 0.3M Na2HPO.7H2O. The pH value was 2.5. The samples were stained for 5 minutes, and rinsed in distilled water. Each slide was observed by a fluorescence microscope using a 490-nm excitation filter and a 530 nm barrier filter on the same day. One hundred cells were counted on each slide. Spermatozoa with normal DNA content exhibited green fluorescence of the heads, while those with denaturated DNA exhibited a spectrum varying from yellow green to red (19). Statistical analysis Paired t-test was used for the comparison of the mean intensity of the sperm heads reactions before and after freezing and thawing process. p- value of less than 0.05 was considered statistically significant. The analyses were performed by SPSS version 11.5 for windows. Results The sperms reacted with all lectins, however the intensity of the reactions were different. DBA showed the more intense reaction and PNA showed less than the others. The extent of binding of WGA and PNA over the acrosomal region was greater than over nonacrosomal regions, indicating the more concentrate of sialic acid, N-acetylgalacosamin and galactose/ N acetyl galactoseamine (GAL/GALNAC) residues in this area of the plasma membrane. Acrosomal regions as well as middle piece of some sperms were also stained with WGA (Figure 1). Only DBA bound consistently to the sperm surface (Figure 2). The WGA and PNA bound only with variable intensity in the total population of sperms. Statistical analyses revealed that there were significant differences in intensities of the reactions to the various lectins before and after freezing and thawing with p-value less than 0.05. The results indicated that the reactions of 46.67%, 34.09% and 73.34% of the specimens modified to PNA, WGA and DBA lectins, respectively, after freezing and thawing (Table I). The modification in the uptake of lectins by sperms of the different specimens was not similar. As the table I shows, some specimens stained more intense while the others uptook less similar lectin after freezing and thawing. Figure 2 depicted the difference between the intensity of the staining of sperms before and after freezing. When the intensity of the reactions of all samples to each lectin were collected and analyzed statistically, it revealed that the intensity of the reactions for DBA and PNA were different with p-value<0.001 (Figure 3). The sperms that stained with acridine orange reveled DNA denaturation. The percents of the sperm with denatured DNA were calculated before and after freezing. Statistical analyses showed that just 4.45% of the specimens contained more sperms with denaturate DNA after freezing and thawing. When the data from all specimens were collected and analyzed statistically, the percentage of the sperms with DNA denaturate nuclei that assessed before and after freezing were 8% and 11% respectively (p- value= 0.058). There was no correlation between the denaturated state of the sperm DNA and modification of the surface glycoconjugates. Discussion Glycoconjugates play important roles in the interaction of the sperm and zona pellucida and also in fertilization (20). Sperm glycocalyx modifications were observed in some kinds of infertility (6). Therefore, any changes in the surface glycoconjugate pattern may potentially cause subfertility or diminish the pregnancy outcome (6). In assisted reproductive technology program, patterns of sperm surface glycoconjugates may change by physical damage during cryopreservation. Therefore, glycol-conjugates modification may lead to change in sperm biological function. The data showed that the acrosomal regions of the sperms head reacted with WGA. The previous study was also shown that WGA binding was very strong on the acrosomal region, weak on the postacrosomal region and moderate on the tail in bull sperms (21). Our observations confirm these data. The data revealed that WGA binding site modified in 34.09% of all samples and when the total samples were analyzed, no difference was observed. Our data showed that DBA binding site were largely restricted to the head region. This data is similar to the other works on lectin binding pattern in mouse sperm (18). DBA binding sites were also detected on human sperms (22). There is also evidence that show the DBA had no distinct staining on spermatogenesis lineage (23). The percentage of the specimens that showed modification N-acetylgalactoseamine was higher than those with the other terminal sugars after freezing and thawing. Carbohydrate is being increasingly recognized for its importance in mediating the adhesion between mammalian sperm and zona pellucida (15). During capacitation, the lectin binding pattern and glycoconjugates distribution change in human sperms (14). It has been also shown that the cross link between certain lectins and their specific sugar motifs influence some sperm characteristics and function such as sperm membrane permeability and acrosomal reaction (3). The terminal charbohydrates that bind to WGA and PNA have important roles in sperm capacitiation (14), permiabilization and acrosomal reaction (3). Modification in PNA binding site after thawing may damage capacitation and acrosomal reaction. PNA can react with components of the acrosomal matrix (24). Acridine orange technique detected single strand DNA in apoptotic cells (25). There is some evidence indicating glycoconjugates modification during apoptosis. One of these glycoconjugates that modify in apoptotic cells is sialic acid which labels with WGA (26). Our data shows that there is no correlation between the DNA denaturation and glycoconjugates modification after freezing and thawing, therefore, it seems that change in lectin binding pattern we observed after thawing is not due to DNA denaturation that may be related to apoptosis in sperms. In conclusion, this is the first work that performed on the effects of cryopreservation on sperm glycocalyx composition. Crystal formation or osmotic changes during freezing may damage surface glycocalyx physically. Chemicals such as DMSO in freezing medium may also have detrimental effect on surface molecules (27) and lead to dissociate the terminal sugars and by this way, it changes the carbohydrate composition by exposing penaltomere sugars. The induction of spontaneous acrosomal reaction may lead to expose the posterior membrane of acrosome with different carbohydrate composition (28-30). These phenomena exert various intensities in each individual sperm and cause the sperms react inconsistently after cryopreservation procedure. Therefore, cryopreservation modifies the surface glycoconjugates pattern and it may have some detrimental impact on fertility of sperm that use in assisted reproductive technology. Acknowledgment The authors wish to thank the research deputy of Shiraz University of Medical Sciences for offering the grant no 2839. References
© Copyright 2010 - Iranian Journal of Reproductive Medicine The following images related to this document are available:Photo images[rm10021f1.jpg] [rm10021f3.jpg] [rm10021t1.jpg] [rm10021f2a-b.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}