 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
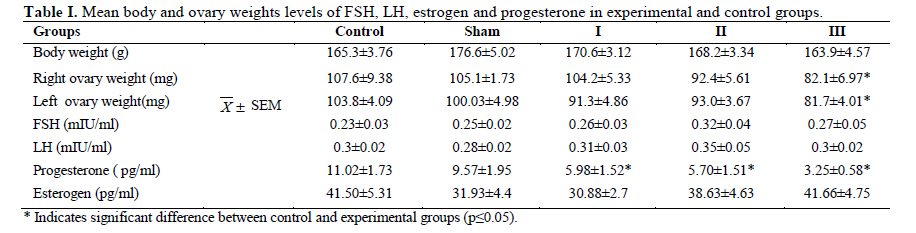
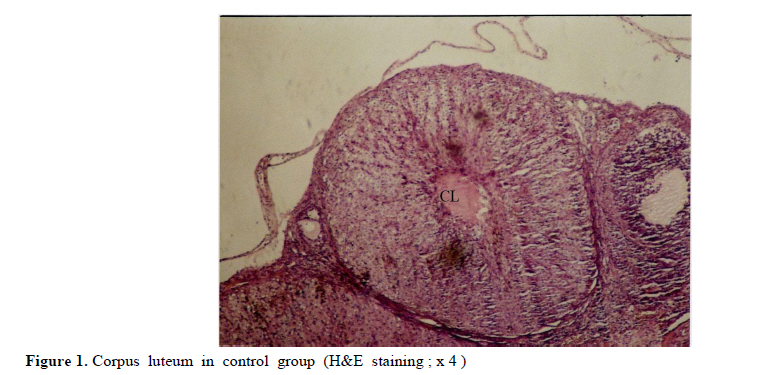
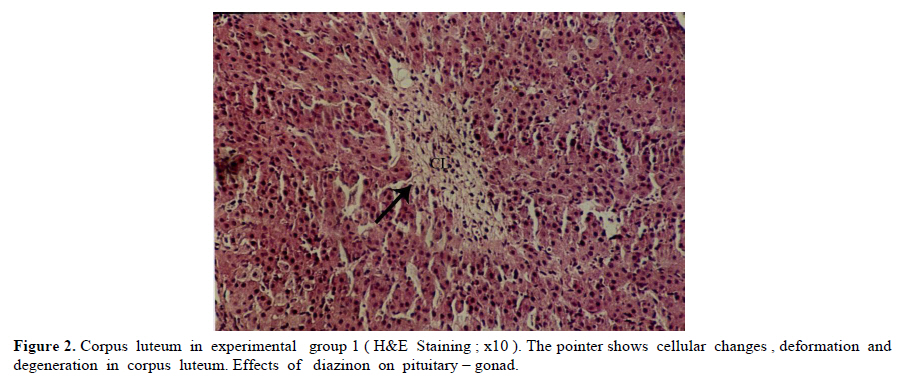
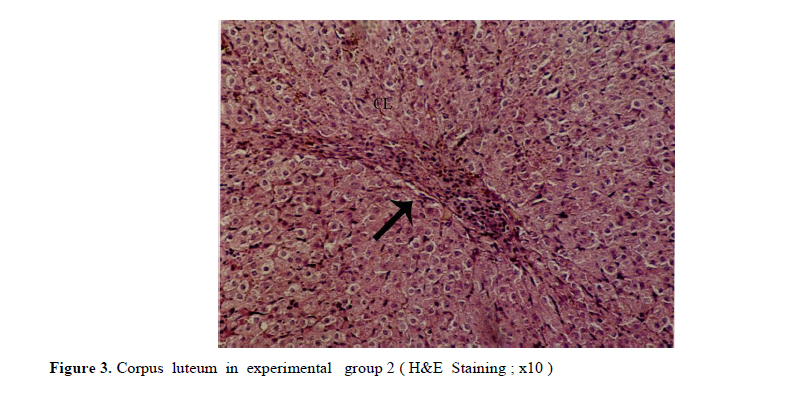
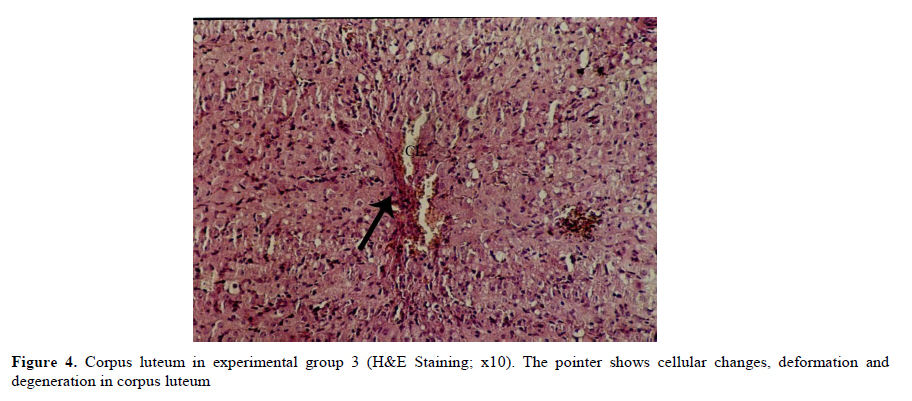
Iranian Journal of Reproductive Medicine Vol. 8, No. 3, Summer, 2010, pp. 125-130 The effects of diazinon on pituitary–gonad axis and ovarian histological changes in rats Habibollah Johari1 Ph.D., Mehrdad Shariati2 Ph.D., Shahram Abbasi2 M.Sc., Esfandyar Sharifi2 M.Sc., Hamid Reza Askari2 M.Sc. 1Department of Biology, Islamic Azad University, Darab Branch, Darab, Iran. Corresponding Author: Habibollah Johari, Department of Biology , Islamic Azad University of Darab , Islamic Azad University Avenue , P.O.BOX : 74817 – 83143 , Darab , Iran . Email: johari@Iaudarab.ac.ir Received: 11 October 2009; accepted: 26 April 2010 Code Number: rm10022 Abstract Background: Diazinon is a widely used Organophosphate insecticide, which is applied against plant pests. This compound has various side effects because it acts as an acetyl cholinesterase enzyme inhibitor. Key words: Diazinon, Progesterone, Estrogen, Ovary, Rat. Introduction Exposure to pesticides may be a major cause of various disorders and reproductive dysfunction in humans and animals. Organophosphates are a group of widely used insecticides in agriculture. A German chemist investigated the efficiency of organophosphate as insecticides (1). One of this insecticide family is an acid phosphoric esters called diazinon. This compound is a colourless fluid, with the boiling temperature of 84˚C, 12 hours half life (in animals body) and 300 mg/kg lethal dose in rats (2, 3). Diazinon has acquired a wide usage in farms and households against noxious insects. Organophosphate insecticides are considered only slightly toxic to vertebrates. However there are evidences of greater effects on reproductive function (4). In animals, the absorption of diazinon takes place mainly through digestive system, skin and inhalation; its adverse effects are exerted through the inhibition of acetyl cholinesterase in neurons. Toxic symptoms of diazinon in human include: diarrhea, nauseam excessive salivary secretion, agitation and alertness (5). Degradation and excretion of diazinon occur in liver and kidney by the oxidation and hydrolysis: its oxidation is carried out by p450 enzyme, and its excretion is done rapidly; in rats for example, 50% is excreted in urine in 12 hours (6). The results of some studies Indicate that diazinon and malathion used in commercial formulation could be toxic to in vitro fertilization and embryo development (3). Similarly, the insecticide methyl is a reproductive toxicant in the male rat and causes deterioration in the structural integrity of the reproductive organs and also the biochemical parameters in the epididymis (7). A research in 2009 have showed that diazinon treatment of male mice for 30 days brought about significant reduction in sperm counts and spermatogenesis, Leydig and sertoli cells and a decrease in serum testosterone concentration (8). In addition, diazinon can induce testicular cancer and cause a decline in sperm density (9). Some studies have demonstrated that diazinon increases the risk of cancer incidence, especially breast cancer in female farmers (10). It also causes deformation in sperm chromatin, morphology and viability (11). On the other hand, a research on Chinese hamster showed that organophosphates induces sister chromatid exchange (12). Since diazinon has adverse effects on various organ systems specially reproductive system, and since a relatively high percentage of female farmers are exposed to diazinon directly, we tried to investigate the effects of this compound on ovary and female sex hormones in order to highlight its probable undesirable influences and increase awareness in those at risk and thereby exerting more regulatory and control procedures on the usage of this insecticide. Materials and methods Fifty adult female rats with the average weight of 165± 5 grams, obtained from animals’ house of Islamic Azad University of Kazeroon, were randomly divided into 5 groups of 10 animals each: a control, a sham and three experimental groups (I, II and III). All groups were kept separately in special polycarbonate cages under the same environmental conditions of 22-25°C and 12h lights and dark cycle with unlimited access to water and dry rat food. The experimental groups I, II and III received 50, 100 and 150 milligrams diazinon per kilograms live weight per day (mg/kg/day) respectively. The control group was left untreated and the sham received equivalent amount of distilled water as solvent. For 14 days, diazinon was administered orally via syringes equipped with feeder. Before the start of the experiment, all the rats were sexually co-cycled in a lab lacking male rats; co-cycling was brought about by injection of 100 micrograms steradiol valerte dissolved in 0.2 milliliter olive oil and after 42 hours 500 micrograms progesterone dissolved in 0.2 milliliter olive oil was also injected. To ascertain the co-cycling vaginal smear was prepared from all animals. To prepare vaginal smear vagina was washed with physiologic serum several times, and was spread on a slide. The slides were examined under light microscope after Gimmsa staining (13, 14). Twenty four hours after the last treatment, all the rats were weighted, anesthetized by ether and blood samples were taken from the heart directly. In order to separate sera, blood samples were centrifuged at the rate of 5000 RPM for 10 min; serum levels of LH, FSH, estrogen and progesterone were determined by radioimmunoassay method using special rat kits (Specteria , Finland ) ( 15, 16 ). For histological studies, ovaries were removed, weighted, and placed in Boin ,s solution and after dehydration stepwise in increasing ethanol samples were cleared by xylene and undergone routine histological processes . 5 µm sections were then prepared and stained by Hematoxilin–Eosine dyes. Primary, secondary and graffian follicles, and corpus luteum in each group were counted (13, 14, 17). Statistical analysis The data were statistically analyzed using SPSS software; mean concentrations of hormones and the ovarian cell counts of various groups were compared with the control using the ANOVA and t-test methods, and p<0.05 was considered as a significant difference. Results There were no significant differences in the body weight among various groups; except in experimental group III (dosage of 150 mg/kg) which showed a decrease in body weight (Table I). In addition, the results of hormones assay, demonstrated that the serum concentrations of LH, FSH and estrogen comparable in experimental groups; while the progesterone hormone level decreased significantly in all experimental groups (p<0.05) (Table II). Histological studies of the ovarian tissues showed no significant difference in the mean number of primary, secondary and graffian follicles but there was a significant decline in the mean number of corpus luteum in experimental group 3 (Tables III and IV). Also, we found signs of degeneration in corpus luteum (Figures 1, 2, 3, 4). Discussion Organophosphates affect mammalian brain development through a variety of mechanisms beyond their shared property of cholinesterase inhibition. The idea is that, different organophosphates target multiple pathways (involved in neural cell development) and they deviate in key aspects that may contribute to disparate neurodevelopmental outcomes (18). Several studies show that organophosphates pesticides exert several effects on mammalian spermatogenesis (8). Nevertheless, their effects on oogenesis are less studied. After 24-h exposure to 250 microM malathion or 900 nM diazinon, oocytes survival was significantly decreased, and the negative effect of these insecticides on the regulation of genes encoding proteins involved in transcription (BP75), translation (ribosomal protein S5), and mitochondrial function (cytochrome oxidase subunits I and III), provide evidence for OP insecticides as toxicants for mammals oocytes during the early oogenesis (19). According to our finding, diazinon has no effect on body weight. In 2003, it was demonstrated that feeding baboon with low concentrations of diazinon for 26 months caused no significant changes in body weight, and concentrations of serum lipids and cholesterol lipoprotein (20). In 1980, the effects of 0.1-15 ppm diazinon on rats were investigated for 92 days, and it was shown to have no influence on diet (21). Our study showed that diazinon significantly decreases ovarian weight and mean number of corpus luteum. Like other Organophosphates, diazinon can increase ACTH; this may promote protein catabolism in ovary which could lead to ovarian weight lost (21). Our investigation shows that diazinon decreases progesterone level but it doesn't induce significant changes in gonadotropins and estrogen concentrations. According to different studies, organophosphates significantly reduce plasma level of progesterone (23), by the possible mechanism of corticosterones elevation that lead to a decrease in 3β-HSD activity which is necessary for progesterone generation (24). Most cholesterol used in steriodogenesis, originate from intracellular sources; indeed, this is the restricting step in steroidogenesis; in fact, a few proteins like Steroidogenic active poly peptide (SAP) and Peripheral Benzo diazpins receptor (PBR) act as cholesterol regulators. However, the protein most studied in this respect is STAR. Most organophosphates are able to interfere in STAR protein gene expression, and reduce its level; thereby, decrease progesterone generation (25, 26). According to some studies, prolactin secretion is controlled by dopamine. On the other hand, some organophosphates, like Dichlorvos, stimulate dopamine level and hence, lead to a decrease in prolactin concentration. Since prolactin can elevate the number of LH receptors, its decrease can reduce the quantity of LH receptors which are involved in progesterone synthesis (27 and 28). Some organophosphates inhibit G-protein activities and could lead to inactivation of LH receptors (29); hence, it may reduce progesterone level. Our histological studies show signs of apoptosis in corpus luteum of experimental groups. According to some reports, diffusion of ca++ could activate caspase 12 located on endoplasmic reticulum (30). Likewise, diazinon and other organophosphates induce ca++ release from intracellular structures and promote caspase 12 activation; this could be the possible mechanism for apoptosis incidence in corpus luteum induced by diazinon (31). On the other hand, diazinon can cause apoptosis through mechanism of ATP decline and thus, bring about changes in mitochondrial membrane potential (32). Some organophosphates activate caspase 3 (33), which is another important apoptotic enzyme. Diazinon can accelerate the synthesis of Interleukin II which in turn, elevate prostaglandins; thereby, causing corpus luteum degeneration (34). Prolactin is an important hormone involved in protection of corpus luteum in many species. However, organophosphates decrease proloctin (35, 36) and through prolactin reduction, they could shorten corpus luteum lifetime. Conclusion Our data showed that, 14 days application of diazinon insecticide can cause a reduction in serum progesterone concentration and ovary degeneration; therefore, it is strongly recommended that protective measures should be considered before application of diazinon. Acknowledgement The authors wish to thanks the research office of the Islamic Azad University of Kazeroun, Iran for its financial supports. References
© Copyright 2010 - Iranian Journal of Reproductive Medicine The following images related to this document are available:Photo images[rm10022f4.jpg] [rm10022f3.jpg] [rm10022t2.jpg] [rm10022t1.jpg] [rm10022f2.jpg] [rm10022f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}