 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Iranian Journal of Reproductive Medicine, Vol. 8, No. 4, 2010, pp. 145-152 Co-expression of galectin-3 and αvβ3 integrin at pinopodes of human endometrium Hossein Nikzad1*Ph.D., Maryam Kabir-Salmani2,3* Ph.D., Shigetatsu Shiokawa4 M.D., Ph.D., Yoshiro Akimoto5 Ph.D., Mitsutoshi Iwashita4 M.D., Ph.D.
Corresponding Author: Hossein Nikzad, Department of Anatomy, Anatomical Sciences Research Center, Faculty of Medicine, Kashan Medical Sciences University, Kashan, Iran. Email: hnikzad@yahoo.com *These authors have contributed equally to this work. Received: 3 December 2009; accepted: 20 May 2010 Code Number: rm10025 Abstract Background: Pinopodes are suggested as biological markers of uterine receptivity,
but their molecular components are unknown. Key words: αvβ3 integrin, Endometrium, Galectin-3, Human, Implantation, Pinopode. Introduction Human endometrium is responsive to ovarian steroids and consequently undergoes morphological and biochemical changes during the course of menstrual cycle in preparation for embryo implantation (1). Uterine epithelium is receptive to the blastocyst only during a very short period of time in the luteal phase, so called implantation window (2). In this regard, pinopode formation and αvβ3 integrin expression are the two most cited biomarkers postulated to frame the window of implantation (3). Pinopodes are progesterone-dependent projections of apical membrane of luminal endometrial epithelium, which appear between days 20 and 21 of the natural menstrual cycle and may indicate endometrial receptivity (4). Despite the strong correlation of pinopodes with implantation process (5), the exact mechanisms of pinopodes functions in human are unknown. In rodents, pinopodes are proposed to prevent the cilia from sweeping off the blastocyst, promote withdrawal of uterine fluid and facilitate embryo opposition (6, 7). Regarding the fact that pinopodes of the luminal endometrium are the first areas of contact with floating blastocyst, it is tempting to propose that the adhesion molecules on the pinopodes may have a pivotal role in initial steps of blastocyst attachment to uterine wall. Several adhesion molecules have been proposed to contribute in the attachment of blastocyst to endometrial epithelium, including integrins, lectins and cadherins (8, 9). Previous reports demonstrated that three integrins (αβ1, α1β4, and αvβ3) were expressed in uterine epithelium during the implantation window (10-12). Localization of αvβ3 integrin at the apical plasma membrane of the surface endometrial epithelium (10, 12) and at the apical surface of the implanting embryo (8) gives a significant role to this integrin as mediator of embryo-maternal adhesion. On the other hand, at the morphological level, there are parallels between leukocyte extravasation from the vasculature, in which lectins play central role and the attachment of the human embryo to the uterine wall (13). Lectins provide special mechanisms enabling cell adhesion under flow, which is the requisite first step of adhesion of the free-floating blastocyst stage embryo to the uterus wall (13). Galectin-3, a 31-kDa protein, is an endogenous soluble β–galectoside binding lectin that belongs to an expanding family of animal lectins called galectins (14). This multifunctional lectin is expressed in a variety of tissues and cell types and can be found in the cytoplasm, nucleus, on the cell surface, and in the extracellular space. Lectin is playing a key role in many physiological and pathological processes, as well as implantation (15-20). Due to affinity of galectin-3 for polylactosamine glycans, it binds to glycosylated extracellular matrix components, including laminin, fibronectin, tenascin and Mac-2 binding protein (15). Furthermore, integrins are receptors for galectin-3 (21), and galectin-3 appeared to modulate integrin activation (15, 20). The proposed extracellular role for galectin-3 to bind and cross-link glycoproteins besides its role as a nonintegrin receptor makes it a strong candidate for a mediator of conceptus-endometrium interaction during implantation. To support this, immunoreactive galectin-3 is found selectively localized at the implantation site of mouse embryo, uterine natural killer cells, and several types of placental trophoblast cells whereas it was absent from the uteri of nondecidualized endometrium (22) that implies a pregnancy-related function for this lectin at the maternal-fetal interface. Further, function of galectin-3 in cell growth, differentiation, chemoatraction, adhesion and migration (23), which are critical for initial steps of embryo implantation make it attractive potential participant in the complex events of embryo implantation. However, the involvement of this lectin at implantation sites especially in human is unknown. Thus, this study was designed to examine the expression of galectin-3 in the human endometrium particularly in pinopodes and to detect its co-expression with αvβ3 integrin at the time of implantation window. Materials and methods Endometrial specimens In this prospective clinical study, endometrial biopsies were obtained from the anterior wall of the uterine cavity of 12 women, by using a Randell Curette (Stille Werner). The ethics committee of Kyorin University approved the design of the study and informed consent was obtained from all participating women. All women were fertile with regular menstrual periods (25-35 days). The mean age was 37 years (range 25–45) and none of them had used steroidal contraceptive or an intrauterine device for at least 3 month before sampling. For endometrial dating, according to the histopathological criteria of Noyes (24), the paraffin-embedded biopsies were stained with hematoxylin and eosin and evaluated by an experienced observer who was blind to the study. The specimens that showed pinopodes and were in phase and showed no pathological feature were divided into two experimental groups: early luteal (days 17-19) and mid-luteal (days 20-24). Each biopsy was divided into three pieces; one was fixed in 10% neutrally buffered formaldehyde for immunofluorescent microscopy. The second portion was fixed in 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4) for immunogold electron microscopy and the third portion was fixed in 2.5% glutaraldehyde in 0.1 M phosphate buffer (pH 7.4) for scanning electron microscopy. Scanning electron microscopy
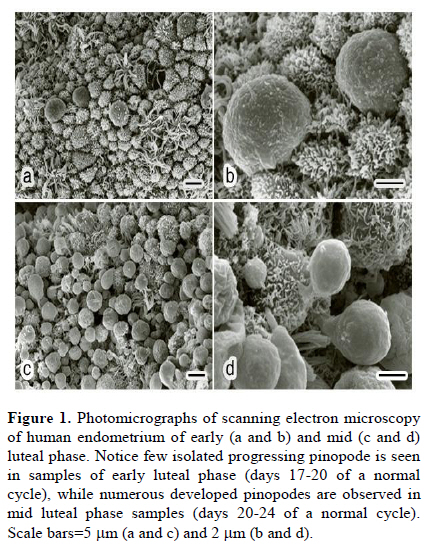
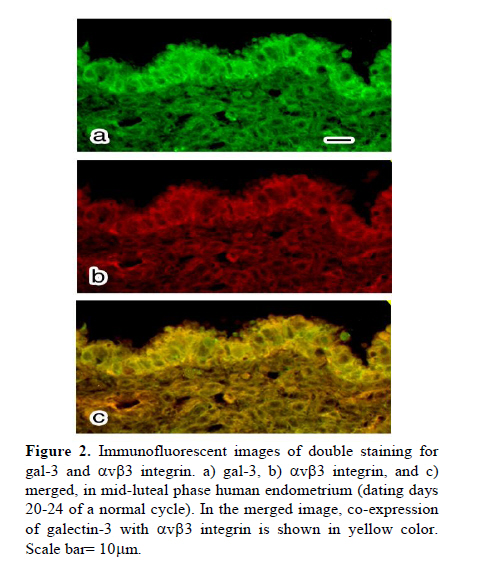
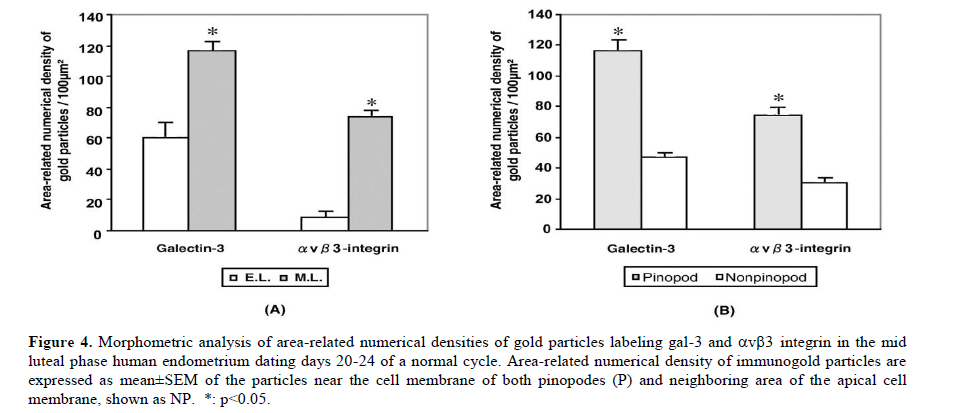
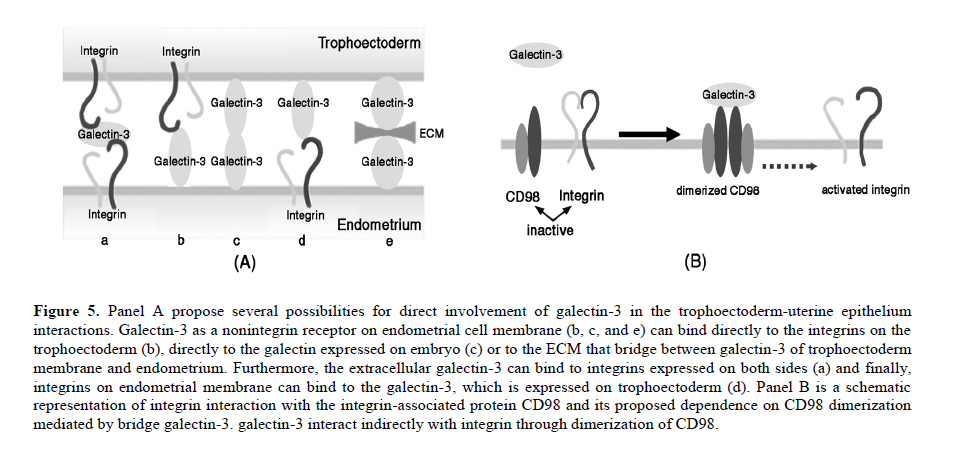
Scanning electron microscopy was used in this study to confirm the presence of pinopodes in the endometrial samples. For scanning electron microscopy preparation, endometrial tissues were fixed for at least 24 hr in 2.5% glutaraldehyde in 0.1 M phosphate buffer (pH 7.4) and post fixed for 1hr in 1% osmium tetroxide in 0.1 M phosphate buffer (pH 7.4). The samples were dehydrated in a graded series of ethanol (50%, 70%, 90%, 99.5% and 100%), critical-point-dried with carbon dioxide by using a freeze drying device (JFD–300, JEOL, Tokyo, Japan), mounted and coated with gold in a sputter coater (JFC-1300 Auto Fine Coater, JEOL, Tokyo, Japan). Finally, the samples examined by using a scanning electron microscope (JSM-5600 LV SEM, JEOL, Tokyo, Japan). Immunofluorescent staining Double staining for galectin-3 and αvβ3 integrin in mid-luteal phase endometrium was performed using immunofluorescent technique to detect their co-expression. For immunofluorescent study, the paraffin sections were processed as mentioned above for DAB staining and blocked by 5% BSA. The sections were then incubated over night at 4˚C with the appropriate primary antibodies diluted in PBS (anti-galectin-3 IgG 1:100 and anti-αvβ3 integrin IgG 1:100). For control, the sections were incubated over night at 4˚C with the same concentration of mentioned antibodies, normal rabbit serum (substituted for anti-galectin-3) and normal mouse serum (substituted for mouse anti-αvβ3 integrin) primary antibodies. The sections were rinsed in PBS extensively and counter-stained with proper fluorescent-labeled secondary antibodies (Alexa 488-labeled donkey anti-rabbit IgG 1:250 and Alexa 568-labeled goat anti-mouse IgG 1:250) appropriately and incubated for 1 hr at room temperature. After washing with PBS, rinsing in deionized water and mounting, sections were observed using an AX-80 fluorescence microscope (Olympus Optical, Tokyo, Japan). These experiments were repeated four times in different endometrial samples taken from mid luteal phase. Immunostaining for electron microscopy Immunogold labeling was performed to quantify ultrastructural distribution of galectin-3 and αvβ3 integrin molecules according to the previous reports (25, 26). Briefly, specimens were divided into 2 mm3 blocks and fixed in 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4) for at least 24 hr at 4˚C. After dehydration in a graded series of ethanol (50%, 70%, 90%, 99.5% and 100%), they were embedded in Lowicryle White Resin (London Resin company Ltd, London, UK) and ultrathin sections were cut. Then ultrathin sections were washed with PBS and pretreated with 5% BSA for 10 min at room temperature. After a PBS rinse, they were incubated with rabbit polyclonal galectin-3 (1:100 dilution) and mouse anti-αvβ3 integrin IgG (1:100) or with normal rabbit and mouse serum as control for overnight at 4˚C. Following washing with PBS 5 times (5 min each), the sections were incubated with the colloidal gold-conjugated (18 nm in diameter) donkey anti rabbit IgG and (12 nm) goat anti-mouse colloidal gold-conjugated IgG (Jackson Immuno Research Laboratories Inc., West Grove, PA, USA) for 1 hr at room temperature (1:20 dilution). Then, the sections were washed with PBS, 5 times and then with distilled water 3 times (5 min each). The ultrathin sections were stained with uranyle acetate and then examined with a transmission electron microscope (JEM-1010; JEOL, Tokyo, Japan). Six different samples from each experimental group were examined using above technique. To detect the developmental changes on expression of galectin-3 and αvβ3 integrin in the human endometrium from mid luteal phase dating days 20-24 and early luteal phase dating days 17-20 of a normal cycle, morphometric analysis was performed. For morphometric analysis of immunogold-labeled galectin-3 and αvβ3 integrin on pinopodes, 240 fields (each field equals to 8.04μm2) per test group (40 fields from each biopsy) were randomly chosen near the cell membrane of the pinopodes by an observer who was blind to the identity of these grids and the number of immunogold particles was counted in the same magnification (X15000). Then, the area-related numerical density of the immunogold particles was calculated. In another set of experiment, to evaluate the significance of expression of galectin-3 and αvβ3 integrin in the pinopodes, in the biopsies from mid luteal phase, area-related numerical density of immunogold particles near the cell membrane of both membrane projections (pinopodes) and neighboring area of the apical cell membrane (not showing pinopode) was evaluated similar to the method that is mentioned above. Statistical analysisIn morphometric assessment of area-related numerical density of immunogold-labeled galectin-3 and αvβ3 integrin, statistical analysis was calculated by averaging a mean of immunogold particles in 40 fields per block for each specimen. Area-related numerical density of immunogold particles were expressed as mean±SEM. Statistical significance were evaluated using ANOVA with Scheffe’s test and were considered statistically significant if p< 0.05. Results Images from scanning electron microscopy demonstrated that endometrial epithelium in secretory phase shows two different types of cells: ciliated and non-ciliated, that latter cover the majority of luminal surface (Figure 1). Membrane projections on the apical pole of non-ciliated cells appear as fine microvilli and dome-like projections defined as progressing, developed and regressing pinopodes (27). Comparing images from early and mid luteal phase specimens revealed that in mid luteal group specimens (dating 20-24) fully developed pinopodes are dominant beside few regressing pinopodes whereas in the specimens obtained in the early luteal phase (dating 17-19) progressing and few isolated developed pinopodes could be detected (Figure 1). In immunofluorescent double staining, co-expression galectin-3 with αvβ3 integrin was observed at mid luteal specimens of human endometrium (Figure 2). The expression of these proteins was shown in the both luminal epithelial cells and stroma, the former showing stronger staining. In negative controls no staining were observed (data not shown). In photomicrograph from immunogold transmission electron microscopy, both galectin-3 and αvβ3 integrin were observed over cytoplasm, nucleus and cell membrane of specimens from early and mid luteal phase endometrium. While, no reactivity was observed on the endometrial surface using the negative control antibody or in specimens incubated without primary antibody in any of the specimens (Figure 3). Further, distribution of these proteins over our two experimental groups displayed a different pattern (Figure 3). Statistical analysis of area-related numerical density of immunogold particles revealed that distribution of these proteins in specimens from mid luteal phase was significantly (p< 0.05) higher compared to those seen in early luteal specimens (Figure 4). Moreover, assessment of area-related numerical density of immunogold particles over the area near the cell membrane of pinopodes in comparison with neighboring area of cell membrane that is pinopode free displayed a higher density of expression of these proteins in pinopodes (Figure 4). Discussion An interesting and to our knowledge a new finding in this study is the ultrastructural localization of galectin-3 and αvβ3 integrin in the apical membrane projections of human endometrial epithelium, so called pinopodes or uterodomes. Pinopodes on the epithelial surface are visible in light microscopy, but other structures may be mistaken for pinopodes (28) and it is not possible to state anything about their stage and the molecular property of these organelles with these techniques. Thus, scanning electron microscopy was used in this study to confirm the presence of pinopodes in the endometrial tissue. Further, using the indirect immunogold technique in this study permitted to evaluate the subcellular distribution of galectin-3 and αvβ3 integrin specifically in pinopodes just prior and during opening of the implantation window. Previous studies reported that galectin-3 was absent from the uteri of non pregnant endometrium in mouse (22) while the results of immunofluorescent staining and electron microscopy in this study demonstrated that galectin-3 is expressed in early and mid luteal phase human normal endometrium. This finding is somehow in consistent with previous study that reported increase of galectin-3 expression in neoplastic endometrial tissue of human (29). Since initial steps of embryo implantation in human have been characterized as an inflammatory response (30), in which galectin-3 act as a mediator of leukocyte adhesion to endothelial cells (31), the expression of galectin-3 in human endometrium could have a similar role during adhesion of floating blastocyst stage embryo to the uterine wall. As the apical pole of the luminal epithelium particularly pinopodes are the first areas to touch embryo in order to initiate embryo attachment, we detected the ultrastructural localization of galectin-3 in human endometrium. Images obtained from immunogold staining for transmission electron microscopic studies clearly exhibited the existence of this lectin in pinopodes. Since pinopodes are progesterone-dependent organells and the maximal expression of αvβ3 integrin on the human uterine luminal epithelium coincides with the rise in progesterone (32), further we examined the co-expression of galectin-3 with this integrin in endometrium. The results of immunofluorescent staining and immunogold technique in this study demonstrated the co-expression of these proteins in human endometrium at mid luteal phase specifically in pinopodes. Previous studies have shown that αvβ3 integrin is expressed at apical pole of uterine epithelium (10, 11), but this is the first illustration of molecular localization of this integrin to the bulbous ultrastructure of human endometrial epithelium called pinopodes to interact with galectin-3 during the early steps of embryo attachment during implantation. The adhesive function of the galectin-3 has been reported also in other adhesive events of several cell types including, extravasation (33), neurite growth (34) and cancer (16, 35). Morphometric analysis of the results obtained from immunogold transmission electron microscopy displayed an increased in expression of galectin-3 and αvβ3 integrin at uterine pinopodes of mid luteal specimens compared to early luteal phase. Furthermore, results from complementary experiments using immunogold staining revealed that these proteins are distributed with a higher density at area near the cell membrane of pinopodes comparing to similar neighboring areas without pinopode. These findings enhance the significance of pinopode formation in preparation for embryo attachment. As for the increase of expression of αvβ3 integrin, this finding is in agreement with previous reports (10, 11), however, they have roughly compared its expression on epithelium rather than pinopodes. Increase in expression of galectin-3 on pinopodes of mid luteal phase endometrial epithelium enhance its potential role in the initial phase of human embryo implantation. The presence of galectin-3 in the endometrium close to the implantation site (pinopodes of surface epithelium) and its increase during the implantation window (luteal phase specimens) propose it as a biomarker of endometrial receptivity. Since the phenomena of implantation and trophoblast invasion are currently considered as the major limiting factor for the establishment of pregnancy (36) thus, the identification of biological markers of endometrial receptivity may have a prominent clinical significance, by improving implantation rates in assisted reproductive techniques (ART) or by promoting efficient contraception. Whether the expression of galectin-3 may change in infertile women or change in expression of galectin-3 may affect embryo implantation rate are subjects for further in vivo and in vitro studies. Our results together with others are in consistent with one of the proposed scenarios for trophoectoderm-uterine epithelium interactions suggesting adhesion molecules that are expressed on either trophoectoderm or the apical surface of the uterine epithelium will bind to their ligands on the extracellular components of their intercellular space to initiate embryo attachment (32). Accordingly, it is tempting to hypothesize that binding of either embryonic or uterine galectin-3 and αvβ3 integrin to their ligands, which are secreted by both cell types (8), could be considered as one of the possible mechanisms during the initial stages of implantation in human, which is proposed in Figure 5A. In this schematic proposal, galectin-3 as a nonintegrin receptor on endometrial cell membrane can bind directly to the integrins on the embryonic side or to the galectin expressed on embryo. Furthermore, the extracellular galectin-3 can bind to integrins on embryonic side or bridge the integrins expressed on both sides and finally, these molecules may interact indirectly through dimerization of CD98 (Figure 5B). CD98 is a disulfide-linked 125-kDa heterodimeric type II transmembrane glycoprotein, which is an important regulator of integrin-mediated adhesion events (37). It has been reported that galectin-3 is an endogenous cross-linker of the CD98 antigen, leading to the activation of integrin-mediated adhesion (21). To examine the co-existence of CD98 with either galectin-3 or αvβ3 integrin at apical pole of uterine epithelium in human, immunofluorescent doubles staining was performed and showed their co-expression in mid luteal phase specimens (results not shown). Further studies need to illustrate the interaction of this tertiary complex at biological level. Taken together, our findings demonstrated the expression of adhesion molecules such as galectin-3 and αvβ3 integrin on the projections of the apical membrane of the human uterine epithelium at mid luteal phase, the time of implantation. Further, the expression of these molecules has been shown to increase during the implantation window. Temporal and spatial expression of galectin-3 in the human endometrium suggests a role for pinopodes and this lectin in initial phases of embryo-endometrial attachment during human embryo implantation. Acknowledgment Authors like to appreciate M. Fukuda, T. Shibata and S. Matubara (Department of Anatomy, Kyorin University, School of Medicine) for their technical assistance in electron microscopy. This work was supported in part by Grants-in-Aid (M.K-S) from NIGEB, Iran project code: 305. References
© Copyright 2010 - Iranian Journal of Reproductive Medicine The following images related to this document are available:Photo images[rm10025f1a-d.jpg] [rm10025f2a-c.jpg] [rm10025f5a-b.jpg] [rm10025f4a-b.jpg] [rm10025f3a-e.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}