 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Iranian Journal of Environmental Health Science & Engineering,Vol. 3, No. 1, 2006, pp. 19-22 SIMULTANEOUS EFFECTS OF SHAKING AND TEMPERATURE ON VEROTOXIN1 PHAGE INDUCTION FROM VEROTOXIGENIC ESCHERICHIA COLI STRAINS *H. Hosain Zadegan, M. Sattari, M. H. Zahir, A. A. Allame Lorestan University of Medical Sciences, Khoramabad, Iran
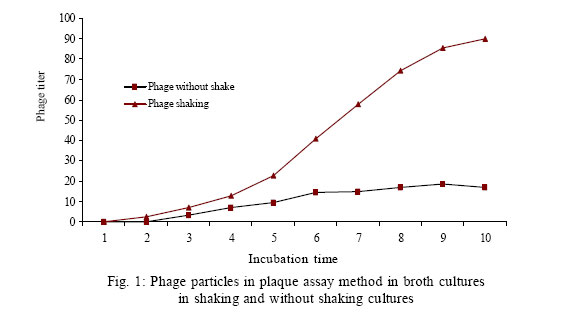
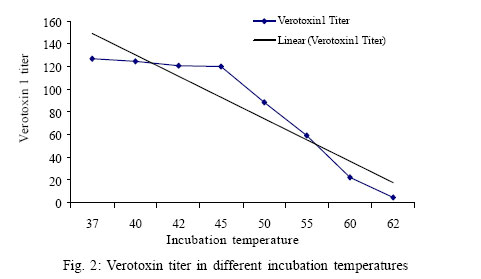
Received 10 July 2005 Code Number: se06004 ABSTRACT Induction of lambda phage carring verotoxin1 gene from a verotoxigenic strains of Escherichia coli and released verotoxin1 were studied under environmental factors of shaking and termperature. Verotoxin1 phage in Escherichia coli PA 101 and transductants was confirmed by bacteriophage detection assay. Shaking of culture media and increasing temperature until 42 ºC increased phage particles in supernatants of Escherichia coli PA 101. Our results indicate that environmental factors such as shaking movements in natural inhabitates of bacteria such as river or sewage streams and temperature rise in summer season could be factors in induce and release free verotoxin1 – producing phage particles in nature that in turn could be the source of phage spreading to other related bacteria , and responsible for increased outbreaks of food borne diseases with verotoxigenic Escherichia coli in warm monthes of year in tropical areas. Key words: Verotoxigenic Escherichia Coli, lambda bacteriophges, verotoxin1 INTRODUCTION Verotoxin producing Escherichia coli (VTEC) are responsible for sporadic cases of diarrhea and have occurring world wide by ingestion of humburgers. (Paton et al., 1998) VTEC causes severe clinical manifestations such as haemorrhagic colitis and haemolytic uremic syndromes. These strains produce shiga like toxins. Verotoxin genes in VTEC are encoded by lambdoid bacteriophage family , that could be induced by factors such as irradiation and mitomycin C. Subinhibitory concentrations of selective antimicrobials for treatment of VTEC outbreaks (Quinolones and macrolides ) also could induce verotoxin genes in VTEC strains. Induction of this genes in turn could be increase verotoxin1 titer in culture or natural environments of bacteria(Yoh et al., 1999 ). Muniesa and Jofre reported that free phages carring verotoxin genes are present in sewage. This phages lysogenized E.coli ATCC (43888, O157: H7, slt) and produced Verotoxin (Muniesa et al.,1998). Muniesa reported that VTEC bacteriophages are more resistant to chlorination and pasteurization, survive better in water than their bacterial hosts. Therfore free living phage particles are potential factors in the spreading and transmission of verotoxin genes among diverse groups of E.coli and related bacteria (Muniesa et al.,1999). Presence of free infectious phage particles in the environment and potential risk of horizontal transmission to other related bacteria, alerts health care authorities and professionals about risks of VTEC caused disease(Schmidt et al., 1999). These facts indicate that in natural habitates some factors may be help to the induction and spreading of phage particles to other bacterial hosts. In this study we investigate the effects of tow environmental factors shaking and temperature on the induction of verotoxin1 bacteriophage from a VTEC. MATERIALS AND METHODS Bacterial strains and cell lines: PA101 is a non- O157 VTEC isolated from a non symptomatic healthy person that produces only verotoxin1, provided by Dr.M. M. Aslani, Pasteur institute of Iran. E.coli DH5α was used for transduction experiments. Vero lines was purchased from cell bank of Pasteur Institute of Iran. Growth media and verotoxin1 measurement PA101 strains was grown in Trypticase Soy Broth. Overnight cultures were washed several times with fresh media. Verotoxin1 Phage was confirmed by plaque forming assay in PA 101 strain.Verotoxin1 concentration in broth or bacterial pellets were determined using a reverse passive latex agglutination (VTEC-RPLA, E.coli verotoxin detection kit; Denka Seiken. Co,Tokyo, Japan) which can differentiate verotoxin1 and verotoxin2 with anti-Vt1 antibody sensitized latex and anti-Vt2 antibody sensitized latex, in accordance with the manufacturers procedure. Peroplasmic toxin extracted by polymyxin B sulphate (0.3 mg/mL final concentration). Cytotoxic activity of crude and affinity purified verotoxin1 tested in a vero cell assay system. Effects of shaking and temperature on induction of bacteriophage and verotoxin1 titer E.coli PA101 was cultured overnight at 37 ºC on tow sets of TSB without shaking ; and TSB at 150 rpm. Verotoxin1 titer and plaque forming units of phage particles in supernatants determined by VTEC-RPLA and bacteriophage detection methods, respectively. In other experiment, we cultured E.coli PA101 overnight at 37 ºC in TSB with 150 rpm. Then broth divided in 15 sections in tubes with 5 mililiter and incubated in 37 ºC , 38 ºC, and two fold increase from 40 to 62 ºC for 10 hours. Then verotoxin1 titer and phage particles were determined as previously mentioned. Determination of plaque forming units Plaque forming unit experiments was done by E.coli DH5α as a recipient host. E.coli DH5α and PA101 were cultured in luria broth and in TSB (150 rpm) at 37 ºC overnight respectively. Liquid culture of PA101 was centrifuged (5000g) at 4ºC, for 10 min. Supernatant sterilized through 0.22 µm pore size filters. Dilutions of filterate was mixed with 0.5 Mcfarland concentaration of E.coli DH5α, and presence of phage was detected by plaque assay on LB soft agar with %0.7 agarose(Maniatis et al., 1982). Verotoxin1 production in transductants of DH5α (TDH5α) was confirmed by VTEC-RPLA Kit in comparison with negative controls of DH5α. RESULTS Kinetic of verotoxin1 production in culture media Time dependent evaluation of verotoxin1 production indicate that in solid TSB,verotoxin1 detected in pellets of bacteria 3 h after bigining of culture. But in TSB without shaking and TSB with 150 rpm detectable verotoxin1 titer measured after 12 and 4 hours of cultures respectively. PA101 growth in TSB (150 rpm) resulted in a 4 fold increase in yield in comparison with TSB without shaking. Shaking of PA101 strongly increased verotoxin1 amount and phage particles in supernatant. We found less than 5 percent of verotoxin1 in supernatant of unshaked TSB cultures of PA101. Similarly plaque assay of filter sterilized supernatants of unshaked TSB showed low levels of phage particles (Fig. 1). Study of temperature effects on verotoxin1 production indicate that in TSB cultures of PA101 at 37 ºC to 55 ºC, verotoxin1 titer increased with elevation of temperature , but decreased in higher than 55 ºC. Low levels of verotoxin1 presence in 62 ºC incubated samples of PA101 culture (Fig. 2) indicates that pasteurized methods of foods such as milk for inactivation of toxins must be reevaluated. Lysigenization of E.coli DH5α Filter sterilized supernatant of E.coli PA101 grown on TSB (150 rpm) was used to transduce E.coli DH5 α. Transduced bacteria (TDHα) produced high titer of verotoxin1 in TSB at 150rpm shaking. verotoxin1 phages in TDHα was confirmed by plaque assay on Luria broth low and top agars. Production of verotoxin1 in TDH α strain evaluated by VTEC-RPLA Kit. Lysates and supernatants of both PA101 and TDHα had same specificity for killing cultured vero cells. Intraperitoneal injection of both supernatants in Balb/c induced paralysis of hind-leg and death in second week of injection. DISCUSSION Virulence genes are frequently encoded in mobile genetic elements, including bacteriophages. This genes are aquired by pathogenic bacteria via lateral transfer of these elements. (Acheson et al.,1998) Our studies indicate that verotoxin1 bacteriophage could be easily transfer horizontally to other related bacteria, and environmental factors such as shaking of contaminated material and temperature fluctuations could also help to releasing and inducing verotoxin1 phage in natural habitates. Kohler reported that lambdoid phages of E.coli in vivo induced under poor environmental conditions such as starvation,aggressive chemicals, and UV light. (Kohler et al.,2000) Muniesa reported that verotoxin1 producing bacteriophages survive better in water than their bacterial hosts and are more resistant to chlorination and pasteurization. (Muniesa et al., 1998) Kohler reported that subinhibitory concentrations of the antibacterials growth promoters of quinoxaline-1,4 –dioxide- type antibiotics olquindox and carbodox increased the release of STX-converting phage particles from Shiga toxin producing E.coli (Kohler et al., 2000). Kimitt reported that quinolones such as ciprofloxacin, ofloxacin, and nalidixic acid have inducing potentials for verotoxin2 production. (Kimmitt et al., 2000)Acheson reported that SLTcarring phage (H19B) have potential of infecting other serotypes of E.coli (Acheson et al., 1998). Feng reported that coliphages MS2 and Qβ have different inactivation rates in a range of pH and temperature(Feng et al., 1992) Helge indicated that clinical SLT-producing E.coli isolates lost their toxin genes during subcultivation and become nontoxigenic(Karch et al., 1992) Our results in lossing of verotoxin1 gene in broth cultures are in accordance with Helge studies. Kajiura assesed the potential of four free phages carring slt genes from Enterohemmorrhagic E.coli O157:H7 (Kajiura et al., 2001) Ohnishi reported that horizontal gene transfer plays central role in generating genomic diversity among E.coli strains(Ohnishi et al., 2001). Our results indicate that verotoxin1 phage and toxin amount in in vitro increased after shaking and incubation of grown strains in higher temperatures. These findings show that antibacterials and environmental conditions contribute to induction and releasing of verotoxin1 phage in different biological ecosystems, and could help to spreading of verotoxin-producing genes among bacterial hosts and producing new Verotoxin producing E.coli strains. In addition, presence of verotoxin1 in hightemperature incubated broths of E.coli PA101 indicate that fast pasteurization methods for foods such as milk could have risks for public health and necessitates exact quality control for pasteurization processes. REFERENCES
© 2006 Tehran University of Medical Sciences Publications The following images related to this document are available:Photo images[se06004f1.jpg] [se06004f2.jpg] |
| |||||||||

{kind=link}
{kind=link}