 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Iranian Journal of Environmental Health Science & Engineering,Vol. 3, No. 4, 2006, pp. 255-260 CONTROL OF ENVIRONMENTAL HEALTH AND REMOVAL OF RATS BY α2-ADRENERGIC ANTAGONIST AND POTASSIUM CHANNEL BLOCKER *1M. Khosravi, 1S. Oryan, 2S.A. Haeri Rohani, 1K. Parivar, 3R. Marandi 1Science and Research Branch, Islamic Azad University, Tehran. Iran *Corresponding author-Email: maryam.khosravi@yahoo.com Tel: +98 21 2294 2275 Received 18 May 2006; revised 10 June 2006; accepted 26 August 2006 Code Number: se06035 ABSTRACT In the present study, it is shown that in vivo oral administration of glibenclamide (1-10 mg/kg/day), an adenosine triphosphate -sensitive potassium channel blocker, and yohimbine (1-5 mg/kg/day), an alpha 2-adrenoceptor antagonist, potently reduced glycemia in male rats. Diazoxide (1-45 mg/kg/day), a potassium channel opener, and clonidine (0.05-0.5 mg/kg/day), an alpha 2- adrenoceptor agonist, antagonized the effects of glibenclamide and yohimbine respectively. Administration of glibenclamide and yohimbine separately decreased the serum glucose levels, so it was expected that coadministration of these two drugs significantly decreased this parameter in fasted animals. It is well known that glibenclamide blocks the potassium channels and it is likely that, yohimbine also blocks these channels, but according to our data, it seems that yohimbine acts via a different binding site to that of glibenclamide. In this research it is also observed that administration of yohimbine and glibenclamide causes death in rats,, it seems that these drugs causes death in the experimental animals by lowering blood glucose level, which were the aim of this study and could be applied as a way to control the environmental health by abolishing rats. Key words: Rats, KATP channel, glibenclamide, yohimbine, environmental health INTRODUCTION In stress, sympathetic system stimulation inhibits insulin release via postsynaptic alpha 2 (α2)-adrenoceptors on the pancreas beta cells, intense sympathetic stimulations interrupt the activation of beta cells in type 2 diabetes mellitus (DM). Some of the researchers believe that pretreatment with clonidine potentiated hyperglycemia. Glycogenolysis and gluconeogenesis or decrease of glucose intake by tissue cells (increase of growth hormone and epinephrine) may induce the increase of blood glucose (Hiyoshi et al., 1995). In this research, administration of yohimbine (Yo) (á2-adrenoceptor antagonist) separately or combined with glibenclamide (Gl) (adenosine triphosphate (ATP)-sensitive potassium channel (KATP channel) blocker) could decrease blood glucose. By this idea, that administration of Yo and Gl may causes death of rats; oral administration of this drug may be a biologic way to defend against these animals. Archeologic studies have shown that rats were contemporain with human being in the third period of geology. In addition that rats are one of the pests of agricultural products, and produce severe damages to channels, buildings, cable wires and etc. Also data have shown that 35 diseases such as thyfus, peste, bite fever, jaundice, thyphoid, diarrhea, cholera, webbel, pig worm, trishin are transported by rats. The susceptibility of persons is depended to individual resistance, kind and number of microbes. The risk of getting diseases is potentiated by using infected food, because foods are easily infected by waste materials, body hair and urine of the animals. To prevent the propagation of diseases transported by rats, hygienic items must be considered. Foods with animal origin must be cooked very well done. Meal products must be observed very carefully in the butcheries and food stores should be out of the reach of rodents (Ahmadbeygui, 1379). Studies and experiments have shown that a pair of rat (male and female) in one year can produce a family with five hundred members. Cleaning the egoes, environment promoting, physical and chemical fights, electrical and ultra-violet trapping and use of ultrasound are one of the ways to defend against rodents. These models of fights are used to reduce the rat's number and preventing the propagation of diseases common between human and animals easily. Foods containing anticoagulant or toxins can be used for domestic and semi- domestic rodents. But always we should be carefull about the health of human and animals in reach with toxins (Ahmadbeygui, 1379). Chemical toxins are technological products which pollute the environment. In some countries, use of toxins for rats is irradicated and to maintain life environment hygiene, use of these toxins is forbidened. But in Iran these methods are already current and is better that, non-chemical specially biologic methods be used because they can produce a minimal damage for the environment.Natural toxins also can effect with lower doses , can be analysed faster in the nature and can damage a little the environment . At the end, the methods for fighting the rats should be cheap. Chemical toxins are divided into two groups: acute and chronic toxins (Ahmadbeygui, 1379; Ashaghi, 1384). Acute chemical toxins such as Arsenic oxide and strychnine (with high toxicity),Talium sulfate, Aneto, phosphore de zinc (with moderate toxicity), Rodascoil (with low toxicity) chronic chemical toxins such as anticoagulants for example varfarine, comatetricile, chlorate, lanivate and actocine. The mechanism of these toxins is inhibiting prothrombine formation (an anticoagulant factor) (Ahmadbeygui, 1379; Ashaghi, 1384). One of the á2-adrenoceptor selective blocker is yohimbine. Yohimbine is an indole and the principal alkaloid from the bark of the west-african tree Pausinystalia Yohimbe Pierre (Formerly Corynanthe yohimbe) family Rubiaceae (madder family). Yohimbine has many effects on the central nervous system and body organs. This drug has parasympathomimetic effect and can induce parasympathetic activation.The yohimbine effects are anxiety, hypertension, tachycardy, locomotion, erection and convulsion. Glibenclamide is one of the sulfonylureas of second-generation and the blocker of KATP channels. The KATP channel on the beta cell consists of a hetero-octomeric complex of four pore subunits, which are known as inwardly rectifing potassium 6.2 (Kir6.2) , and four regulatory ,or sulfonylurea receptor 1 (SUR1), subunits. These drugs bind to the SUR1 subunit of KATP channel and by blocking these channels, depolarize the cell membrane, opens calcium voltage dependent channels and increase intracellular calcium, then vesicles containing insulin are transported to the cell membrane and secreted out of the cell (Doyle and Egan, 2003; Eliasson, et al., 1997). In the presence of sulfonylureas the activation of KATP channel even in the absence of glucose is stopped and it evokes hypoglycemia in all conditions. When the half-life of sulfonylureas special analogue is longer the possibility of inducing hypoglycemia is more enhanced (Eliasson, et al., 1996). There is evidence that some sulfonylureas actually enhance the usual inhibitory action that MgADP (magnesium-adenosine diphosphate) has on the nucleotide binding site of the Kir6.2 subunit and so this may increase insulin release even more under hypoglycemic conditions. MgADP effects on the KATP channel are 2-fold: it inhibits the Kir6.2 subunit and stimulates the SUR1 activity. However, in the presence of some sulfonylureas, the interaction of MgADP with SUR1is diminished, resulting in its unopposed inhibitory effect on Kir6.2 (Doyle and Egan, 2003). MATERIALS AND METHODS In the present study, normal male wistar rats (170-230 g) purchased from Pasteur institute of Iran, were kept under a 12-h light/dark cycle. Water and food were available, and the room temperature was 21-25 degree of centigrade. Animals were divided into groups of six rats. Drugs used in this study were :
Clonidine and yohimbine are soluble in water. Each day, glibenclamide and diazoxide were prepared in the solvent. The solvent for 30 mg diazoxide was: NaOH 0.9cc (0.1 N). The solvent for 30 mg glibenclamide was: 0.6mL (1M) NaOH + 1mL ethanole + 1mL polyethylenglycol 400 + 0.4mL H2O. Instruments and kits
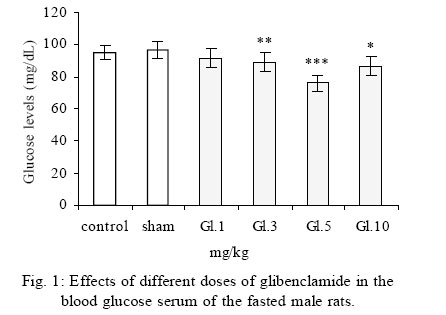
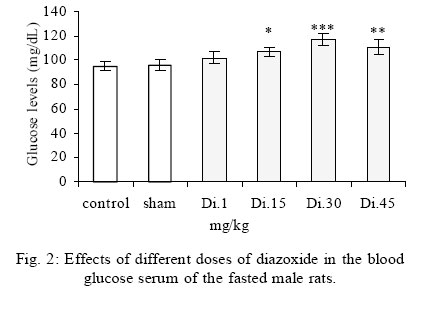
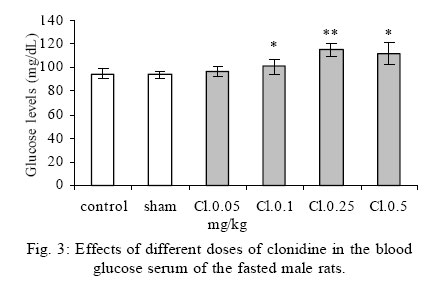
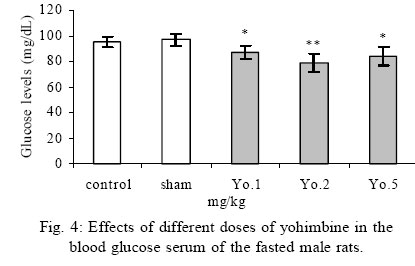
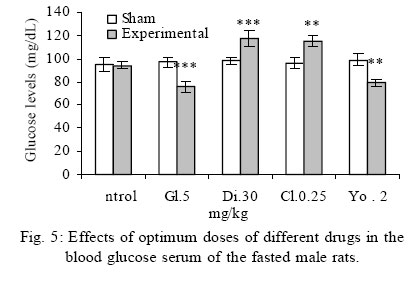
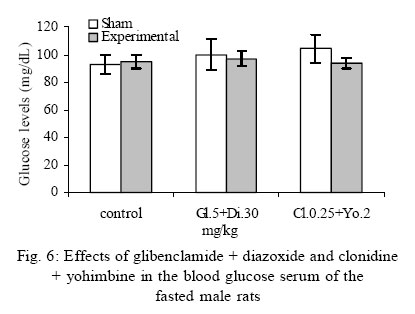
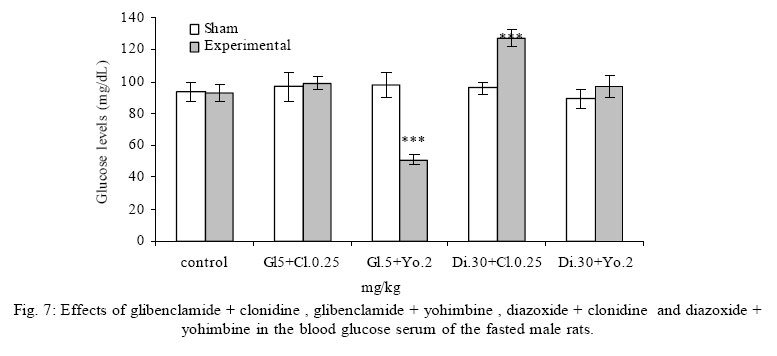
Treatment Drug administration: One night before drug administration, the animals were deprived from food. Then all the groups received oral administration of the drug for one week. In this period of time, water and food were available for the animals including control group. Blood sampling: Rats were anesthetized with ether then blood was sampled from the retro orbital sinus. The blood sample were clotted in the laboratory. Then, the samples were centrifuged for 15 minutes with a speed of 3000/min. Serum glucose level was measured by glucose oxidase (Enzymatic, colorimetric-GOD-PAP, Cat no. 10505 Biochemical). Glucose level was measured with spectrophotometer based on enzymatic method and color intensity with glucose oxidation. Statistical analysis All data were reported as means ±SEM. Differences between groups were examined by one-way analysis of variance (ANOVA) with Tukey test at a significance of *P<0.05, **P<0.01, ***P<0.001. RESULTS Different doses of glibenclamide (K channels blocker) 1, 3, 5, 10 mg/kg administrated to male rats, decrease glucose serum dose dependently and the more effective dose was 5 mg/kg. Different doses of diazoxide (K channels opener) 1, 15, 30, 45 mg/kg increase glucose serum dose dependently and the more effective dose was 30 mg/kg. Clonidine (alpha-2 adrenoreceptor agonist) 0.05, 0.1, 0.25, 0.5 mg/kg increase glucose serum dose dependently and the more effective dose was 0.25 mg/kg , and different doses of yohimbine (alpha-2 adrenoreceptor antagonist) 1, 2, 5 mg/kg decrease glucose serum dose dependently and the more effective dose was 2 mg/kg. In these experiments, it is shown that there is no significant difference of glucose level between control and sham group (intact). But there is significant difference of glucose levels in the groups administrated by glibenclamide (Gl), diazoxide (Di), clonidine (Cl), yohimbine ( Yo ) comparing with their sham groups in Figs. 1, 2, 3, 4, 5. There is no significant difference of glucose level in the groups receiving glibenclamide + diazoxide or clonidine + yohimbine comparing with their sham groups. And also, it is shown that there is no significant difference of glucose level between control and group receiving Gl + Cl or Yo + Di. But there is a significant increase of glucose level comparing control and groups receiving Di and Cl in Figs. 6, 7, and 8. DISCUSSION In this study, glibenclamide (K channels blocker) decreases glucose level dose-dependently. 5 mg/kg of this drug was used as the optimum dose, because in these experiments the lowest glucose serum level was obtained after using this dose of drug. These results are consistent with the results of previous studies (Tian, et al., 1998; Eliasson, et al., 1997). In 1996 it was reported that the action of sulfonylueas may not be entirely limited to closure of the KATP channels, but that they may have a direct effect on the exocytotic machinery of the beta cells. The effect is protein kinase C (PKC)-dependent (Eliasson, et al., 1996 ) but may not involve activation of PKC (Tian, et al.,1998).Renstrom and colleagues have proposed that it is associated with the maintenance in the insulin vesicles of a pH and ion balance conductive to exocytosis (Renstrom, et al.,2002).Acidification of the internal milieu of the insulin vesicle occurs via uptake of H+ and is a necessary priming step for release of insulin. This increase in positive charge within the vesicle must be balanced by an influx of Cl- to prevent an excessive build-up of positive charge and thus permit vesicle acidification. Sulfonylureas are known to bind to a 65-KDa protein found on the vesicle fraction of the beta cell. (Kramer, et al., 1994) which is instrumental in the acidification of the insulin vesicle. Diazoxide also increase blood glucose dose dependently , and the more effective dose of this drug was 30 mg/kg. These results are in agreement with the previous studies (Clapham, et al., 1994). Diazoxide is the more potent opener of KATP channels and its effect are contrast to sulfonylureas. Diazoxide hyperpolarizes beta cells and after inhibiting insulin secretion,cause hyperglycemia .This drug opens KATP channels by the presence of Mg2+ and ATP and it seems that these channels have a different binding site than suffonylureas(Doyle and Egan, 2003). The coadministration of optimum doses of glibenclamide and diazoxide (5,30 mg/kg) show that the hypoglycemia induced by glibenclamide is antagonized with diazoxide.These results agree with the studies of Tian et al., 1998 and demostrate that glibenclamide can block KATP channel and diazoxide is capable to open it. According to the results of the present research, clonidine (alpha-2 adrenoreceptor agonist) increased blood glucose serum dose dependently, and the more effective dose of this drug was 0.25 mg/kg . These results are in agreement with the previous studies of O`Dannell, et al., 1996. It seems that á2-receptors, inhibit adenylyl cyclase activation via the inhibiting regulatory Gi protein (inhibitory G protein). The mechanism which Gi inhibits adenylyl cyclase is not exactly known. But there are two probabilities:
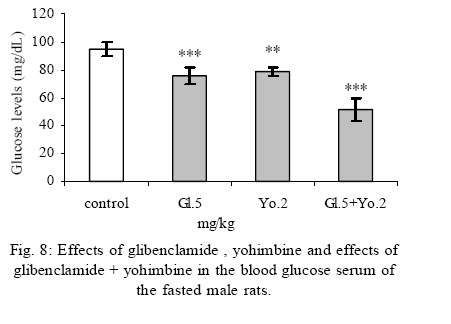
Some researchers accept both of these two mechanisms. In addition , some of the effects of alpha-2 adrenoceptors are not due via adenylyl cyclase inhibiting, these mechanisms include opening of the potassium and closing of the calcium channels .Perhaps these effects are due to the function of the beta-gamma subunits of Gi protein, which are due via ion channels and enzymes. Stimulation of the alpha-2 adrenoceptors in the beta cells is believed to directly activate a G-protein-gated K channel, there by inhibiting exocytosis (Doyle and Egan, 2003).On the other hand, clonidine potentiates the inhibiting effect of adenylyl cyclase on insulin secretion and also it seems that clonidine by glycogenolysis and gluconeogenesis in liver cells causes hyperglycemia. These results show that yohimbine(alpha-2 adrenoceptor antagonist) decreased blood glucose serum dose dependently, and the optimum dose of this drug was obtained 2 mg/kg. These results are in agree with the studies of other researchers(Jonas, et al., 1994, Efanove, et al., 2001). It seems that alpha-2 adrenoceptor antagonists increase insulin release from beta cells of pancreas islets, and probably some of the imidazolines release insulin via closing the KATP channels and voltage dependent K channels. Some of the investigators believe that the binding site of imidazoline is not the same as alpha-2 adrenoceptor in beta cells (Chan, 1998). Some of the researchers believe that imidazoline receptor is one of the part of KATP channel on beta cells (Olmos, et al., 1994).Two mechanisms may have a role in insulin release induced by yohimbine. 1- In low concentration increases insulin which may induce by alpha-2 adrenoceptor blocking. 2- In high concentration increases insulin which may induce by KATP channels blocking (Jonas et al., 1994). The results from effects of coadministration of optimum doses of clonidine and yohimbine (0.25 and 2 mg/kg) show that clonidine limits and blocks blood glucose serum decreasing. So, after inhibiting the effect of yohimbine by clonidine, blood glucose serum approach to the glucose level of sham groups. These results agree with the findings of O`Dannell, et al., 1996. In the present study coadministration of yohimbine and clonidine, prevent hyperglycemia induced by clonidine, so it seems that in addition of previous mechanisms, yohimbine via blocking the postsynaptic alpha-2 adrenoceptor of beta cells has prevented the inhibition of adenylyl cyclase and increased the intracellular cyclic adenosine monophosphate (cAMP). Also the results of coadministration of yohimbine and diazoxide show that yohimbine prevents the hyperglycemia induced by diazoxide. It seems that probably yohimbine like imidazolines binds to a different site of Kir6.2 subunit (subunit forming the channel pore) and blocks the channel. The experimental results in this study show that the coadministration of yohimbine and glibenclamide cause death in rats probably due to the decrease of blood glucose plasma concentration. These drugs are not as harmful and pollutant as the other animal toxins for human being. On the other hand, these drugs are biologic, easily analysed in the environment ,cheap , available and could be easily added to the food or water of the animals. So it is suggested that in the future researches ,the effects of these drugs could be studied as a chemical method to control the pollution of big cities such as Tehran. Adding the supplements to the above mentioned durgs are the future targets of this study and could be extended throughout the country. REFERENCES
© 2006 Tehran University of Medical Sciences Publications The following images related to this document are available:Photo images[se06035f4.jpg] [se06035f3.jpg] [se06035f5.jpg] [se06035f1.jpg] [se06035f7.jpg] [se06035f8.jpg] [se06035f2.jpg] [se06035f6.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}