 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
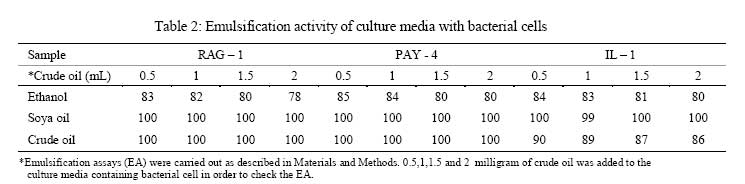
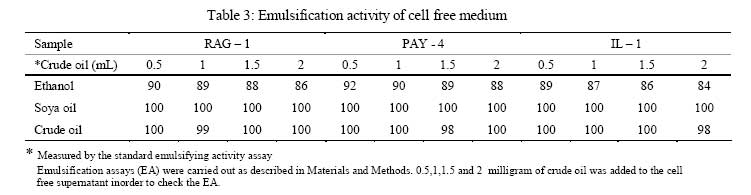
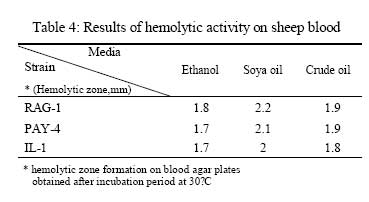
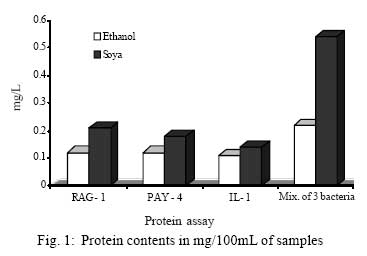
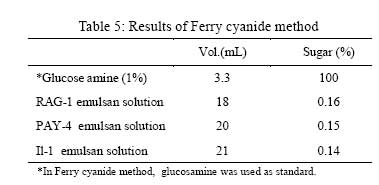
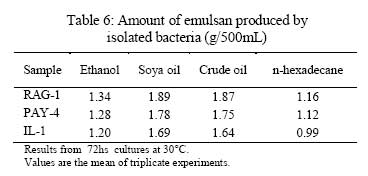
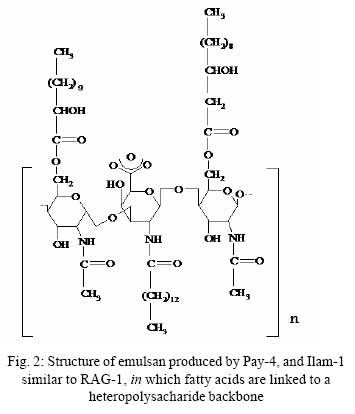
Iranian Journal of Environmental Health Science & Engineering, 2008, Vol. 5, No. 2, pp. 101-108 Emulsan Analysis Produced by Locally Isolated Bacteria and Acinetobacter Calcoaceticus RAG-1 1 P. Chamanrokh, *2 M. Mazaheri Assadi, 1 A. Noohi, 3 S. Yahyai 1 Graduate College of the Basic Sciences, Islamic Azad University, Science and Research Campus, Tehran, Iran Received 23 December 2007; revised 13 February 2008; accepted 4 March 2008 Code Number: se08020 ABSTRACT Growth of previously isolated bacteria from Iranian oil reservoirs on different carbon and energy sources and under varying conditions have been used to produce a class of extracellular microbial protein-associated lipopolysaccharides named emulsan.Several Bacteria were previously isolated from Iranian oil reservoirs and designated as; Ilam-1 and Paydar-4. In present study, the isolated strains were compared with standard sample of Acinetobacter Calcoaceticus RAG-1 from Persian Type Culture Collection (PTCC 1641), IROST. Among the isolated strains, two strains were found to produce an extracellular, emulsifying agent when grown in Mineral Salt Medium containing soya oil, ethanol or local crude oil. The isolated bacteria were cultured and further analysed using protein estimation, reducing sugar analysis, hemolytic activity, surface tension and emulsification activity tests. The crude emulsifier of RAG-1, PAYDAR-4 and ILAM-1 were concentrated from the cell-free culture fluid by ammonium sulfate precipitation to yield 1.89g, 1.78g and 1.69g of bioemulsan respectively. Emulsifying activity was observed over the entire production process. These investigations showed that emulsan produced by isolated Iranian crude oil reservoir were comparable with Acinetobacter Calcoaceticus RAG-1 which is made of carbohydrate backbone as its hydrophilic part (N-acetyl-D-galactoseamine, N-acetylgalactoseamine uronic acid, diamino-6-deoxy-D-glucose) and fatty acid chain as its hydrophobic portion. Key words: Emulsan, emulsification activity, protein estimation, reducing sugar analysis INTRODUCTION Emulsifiers are widely used in industry, e.g., in medical and cosmetic products and in food systems. They can be used as agents to combat oil spills on beaches and in the sea. The materials that are currently in use commercially as emulsifiers are mainly produced by chemical synthesis. There is a great interest in studying bioemulsifiers because they have some advantages, such as their selectivity for specific interfaces, generally low toxicity and biodegradability. The enormous diversity of microbial emulsifiers provides a rich source to find new agents which possess the right combination of properties for specific applications (Rosenberg and Eliora, 1998). A wide variety of petroleum degrading microorganisms has been found to bring about the formation of oil-in-water emulsions while growing on hydrocarbons (Pines, 2005). These emulsans are microbiological in origin and appear to be made either by the cells themselves or by the production of extracellular emulsifying agents (Gutnick et al., 1980). The gram negative bacterium Acinetobacter Calcoaceticus RAG-1 grows on a variety of different substrates as sole source of carbon and energy, including crude oil, middle chain length alkanes, alcohols, fatty acids and triglycerides (Fiechter, 1992). Its molecular weight is about 1000KD in average (Pil, 1997). This molecule is composed of an unbranched polysaccharide backbone with O-acyl and N-acyl bound fatty acid side chains. The polysaccharide backbone consists of three amino sugars, D-galactosamine, D-galactosaminouronic acid and a dideoxydiaminohexose in the ratio of 1:1:1 (Zhang, 1992; Bruce, 2002). The fatty acid side chains range in length from 10 to 22 carbon atoms and can represent from 5% to 23% (wt/wt) of the polymer. The emulsan amino groups are either acetylated or covalently linked by an amide bound to 3-hydroxybutyric acid. The combination of hydrophilic anionic sugar main chain repeated units and the hydrophobic side groups leads to the amphipathic behavior of emulsan and, therefore, its ability to form stable oil-in-water emulsions (Gorkovenko, 1997). Further studies on the microbial degradation of crude oil by this organism (Pines, 2005) showed that RAG-1 emulsified the oil during exponential growth, probably by producing an extra cellular emulsifying agent which acted to break up the oil droplets into smaller units and thereby produce new surface area, necessary for the increasing cell population. At the first International Congress for Bacteriology held on September 2-7 in 1973, Englander reported that this extra cellular emulsifying agent had been partially purified from stationary phase cultures of RAG-1 growing on 0.4% hexadecane, 0.075m urea and 5.8mm dibasic potassium phosphate in sea water. The partially purified extra cellular emulsifying agent was obtained by extensively dialyzing and then lyophilizing the cell-free fermentation broth, yielding 0.25mg/mL of culture fluid of a dry powder which was capable of forming a stable oil-in-water emulsion with 40 times its weight of crude oil (Gutnick et al., 1980). In 2007, there were few publications about this subject; however, microbial induced emulsification of oil still needs clarification from both mechanistic and teleological points of view. Microorganisms can utilize crude oil as a substrate for growth with or without concomitant oil emulsification. Where emulsification has occurred because of the production of extracellular emulsifying agents, in general the preparations have not been purified sufficiently to identify the active components. Although, these extracellular bioemulsifiers have been well characterized but still need analysis on their chemical properties due to broad range of microbial production. Emulsans on a weight for weight basis are probably the most efficient oil-in-water emulsifiers discovered and, moreover, possess certain characteristics that permit these unique extracellular microbial polysaccharides to be widely used in cleaning oil contaminated vessels, oil spill management, and enhanced oil recovery by chemical flooding (Gutnick et al., 1998). To this aim here we report the purification and characterization of produced emulsan and discuss its potential as a biocatalyst in synthesis reactions. MATERIALS AND METHODS Bacterial strain Two bacterial strains previously isolated from Paydar-4 and Ilam-1 reservoir (Amiria et al., 2004), were maintained on Mineral Salt Medium (MSM). After incubation for 3 days, the plates were stored at 4°C. Acinetobacter calcoaceticus Rag1 was obtained from Persian Type Culture Collection (PTCC 1641). Preculture condition To produce the preculture, 1% of bacterial inoculums was used (Acinetobacter calcoaceticus RAG-1, PAY-4 and Ilam -1) in nutrient broth. To obtain the desired growth, flasks were kept for 17 hr in case of IL-1, RAG-1 isolates to reach stationary phase and 19h for PAY-4(OD600=1) (Amirian et al., 2004). Medium and culture condition Bacteria were grown in defined mineral medium which contained (per liter of water): K2HPO4,0.5g; KH2PO4, 2g; MgSO4 7H2O, 0.2g; (NH4)2 SO4, 3g; NaCl 0.1g, 0.03% of the glucose and 0.03% east extract, CaC122H2O, 0.01g; FeSO47H2O, 0.01g and the trace elements: ZnSO4 7H2O, 0.024g CuSO45H2O, 0.25g; MnSO47H2O, 0.17g; CaC122H2O, 0.24g (1mL from trace element was added to the main medium). MgSO4.7H2O was initially prepared as a stock solution (concentrated 100-fold) which was sterilized separately and then added to the culture medium. The seed cultures and fermentations were carried out in 1L. In order to procede for fermentation the baffled shake flasks were used according to the method of Francy, with slight modification (Francy et al., 1991). Each one liter flask contained 200mL of the defined mineral medium. Different carbon sources such as ethanol (2%), soya oil (1%) and crude oil (3%) were used. Size of inoculums From the preculture 3% bacteria was used for the medium (MSM) that contained 7% crude oil. Inoculation of 5% bacteria was used for the medium (MSM) with 1% soya and 3% bacteria was used for the medium (MSM) with 2% ethanol (Nemati and Mazaheri Assadi, 2002). The flasks were placed in a Lab-line incubator-shaker maintained at 30°C (180 and 200rpm). Emulsification activity Emulsification activity was measured using the method described by Francy et al., 1991, whereby (with a modification) 0.5mL of crude oil or other suitable hydrocarbon was added to 2mL of the culture broth in a graduated screw cap test tube and vortexed at high speed for 1min using a vortex-GENIE. The emulsion stability was determined, after 24h. The emulsification index (E24) was calculated by dividing the measured height of the emulsion layer by the total mixture and multiplying by 100. Hemolytic activity on sheep erythrocytes Since hemolytic activity is one of the tools to characterize bacteria, hemolytic activity of bacterial strains was determined by inoculating sheep blood agar; also biosurfactants are metabolites which are capable to destroy the membrane of erythrocytes (Rosenberg and Bayer,1982) and in as a result show hemolytic activity. 2 methods were used: in the first method, emulsan covered disks were put on blood agar plates containing 5, 7, 10 and 12% sheep blood and incubed at 30°C overnight. In the second method, few wells were made in blood agar plates containing 5,7,10 and12% sheep blood. In each well, 70µL emulsan was added. Plates were incubated at 30°C for 48-72h. Clearing zones were checked around the colonies. All experiments were carried out in triplicates (Jain et al., 1991). Lowry protein assay Total protein was determined by the Modified Lowry used to solubilize amphipathic proteins. In this method proteins make a colored complex with Folin reagent. The standard protein curve is drawn for 0, 0.05, 0.1, 0.15, 0.2, 0.25mg/mL concentrations of Bovine Serum Albumin (BSA) in DD.W (Biotek). Also three samples of bacteria, their mixture in soya oil and ethanol were analyzed for protein contents. Determination of reducing sugars For determination of reducing sugars the Modified Ferry cyanide method was used. The solutions of ferry cyanide 1%, sodium hydroxide 2.5N, methylene blue 1% in water were used and used with glucose amine as the standard (Sydney, 1933). Isolation of Emulsan The Emulsan Isolation Method was performed using ammonium sulfate. After the fermentation period, the cultures were centrifuged (8000rpm, 40min), the supernatant was collected, and the pellet was washed once with distilled water. The supernatant and the washing liquid were combined and sulfate (50%) was added to the cell-free liquid which was then maintained at 4°C for 24h. The resulting precipitate was separated by centrifugation (8000rpm, 40min), suspended in water, dialyzed against distilled water for 2 days (at least five replacements of water) and dried by lyophilization. The crude EM was extracted with ether for 2 days using a Soxhlet apparatus and the EM yield was then determined by gravimetric analysis after drying the product in-vacuo (<30 mmHg, 50°C) to constant weight (Gutnick et al., 1980; Rosenberg et al., 1988). RESULTS Biomass from RAG-1, IL-1 and PAY-4 strains Maximum amount of biomass production of three bacteria was achieved after 72h of growth. At 14-36h of growth, the surfactant concentration increased gradually. Maximum production was obtained at about 36h. After 72h of growth, cell dry weights were determined.. The weighed values were 3g/L for A. calcoaceticus and 3.1g/L for PAY-4 and 2.8g/L for IL-1 in crude oil medium. In our previous studies by (Amirian et al., 2004) , more or less similar amount of cell were concluded i.e. 3.4g/L for A. calcoaceticus and 3.2g/L for PAY-4 and 2.9g/L for IL-1 in soya oil medium, 2.9g/L for A. calcoaceticus and 2.8g/L for PAY-4 and 2.7g/L for IL-1 in ethanol medium (Table 1). Culture media and cell free media emulsification activity Emulsification activity values increased with increasing cell growth, reaching their optimum at about 48h and remaining constant until the end of fermentation. Percentage emulsification activity of obtained emulsan with each strain checked, using the modification method of (Francy et al., 1991). Result of this investigation presents experimental emulsifying activities of two isolated whole crude culture media and A. calcoaceticus RAG-1, using 0.5, 1, 1.5, 2mL of crude oil. The emulsion average values (water-in-oil) of about 80.75% was obtained for A. calcoaceticus, whereas 82.25% for PAY-4 and 82% for IL-1 in ethanol media. High emulsion average values (water-in-oil) of about 100% was obtained when soya oil was used as a carbon source with A. calcoaceticus and PAY-4. Comparatively negligible reduction i.e. 99.75% was achieved for IL-1. Emulsion values average (water-in-oil) of about 100% for A. calcoaceticus RAG-1 and PAY-4 and 88% for IL-1in crude oil were comparable(Table 2). In order to find out if there is any difference or interaction of water in oil emulsion when crude was used in culture medium, the product was compared with cell free medium. Samples were centrifuged of 8000rpm for 40min using all the three strains. The maximum emulsan rhamnolipid was produced within 96h. In a resent study we concluded that, soya oil was a good source of carbon for emulsan production. Data in Tables 2 and 3 show the experimental results on the emulsifying activities of the supernatant of two isolates and A. calcoaceticus RAG-1 as standard with 0.5, 1, 1.5 and 2mL crude oil. The emulsion values average (water-in-oil) of about 88.25% for A. calcoaceticus,89.75% for PAY-4 and 86.5% for IL-1 in ethanol media, high emulsion average values (water-in-oil) of about 100% for A. calcoaceticus and PAY-4 and IL-1 in soya oil, emulsion average values (water-in-oil) of about 99.75% for A. calcoaceticus and 99.5% for PAY-4 and IL-1 in crude oil were observed (Table 3). The results showed that there is more or less similarity between the emulsification activity of all the three cultures. These findings suggest that the emulsifier's activity depends on its affinity for hydrocarbon substrates which involves a direct interaction with the hydrocarbon itself rather than an effect on the surface tension of the medium. Emulsification activity values increased with increasing biomass formation, reaching their optimum at about 72h (Table 1). Depicted results of emulsification activity (Table 2 and 3) indicated that produced biosurfactant by two isolated bacteria and RAG-1, had a high emulsification specificity with crude oil and this result differs to that of (Kaplan and Rosenberg, 1982; Abu and Banat,1991); the highest emulsification activity was seen in soya oil media. These findings suggest that the emulsifier's activity depends on its affinity for hydrocarbon substrates which involves a direct interaction with the hydrocarbon itself rather than an effect on the surface tension of the medium. Three types of the biosurfactants remained stable, after exposure to high temperatures of 100oC and 120oC for 15min; similar results were obtained by Kim et al., 2000. Hemolytic activity on sheep blood erythrocytes Biosurfactant producing capacity in liquid medium was found to be associated with hemolytic activity. In soya media RAG-1, PAY-4, IL-1 showed larger zone of hemolytic activity. These zones were 2.2mm for RAG-1, 2.1 for Pay-4, 2mm for IL-1 (Table 4). However, the extent of hemolytic zone formation on blood agar plates is not solely dictated by the concentration of emulsan and may be affected by divalent ions and other hemolytic produced by the microbes. In addition to hemolytic activity of A. calcoaceticus RAG1, other two isolated microbial products lysed blood agar although there were poorly diffusible lysed blood cells. Still it is not clear whether blood agar lysis should be used to screen for bio-emulsan production. Lowry test Protein concentration was compared to bovine serum albumin standards. The role of each isolates in the release of emulsan from the cell surface of all the three strains have been well established. Decline in crude medium protein was recorded to lead to a subsequent rise in the medium and there was an increase in emulsan production too. Using protein standard curve: Bovine serum albumin/DDW (mg/mL) the formula of Y=4.0457X+ 0.031(in which X=protein mg/mL, Y=OD in 650 nm) for measuring the amount of protein. The protein contents were 0.12mg\mL for RAG-1, PAY-4, and 0.11mg/mL for IL-1 and 0.54mg/mL for mixture of 3 bacteria in soya oil medium (Fig. 1). These results suggest that soya oil media showed the highest amount of protein contents rather than other media. A portion of the partially purified bioemulsifier was used for chemical analysis. Ferry cyanide method Production of emulsan macromolecule was measured chemically by the production of reducing groups; approximately 0.016µmol of reducing sugar per mg was generated, corresponding to 0.36% of the glycoside linkages in emulsan being hydrolyzed per hour. The glycoside linkages were broken. After centrifugation to remove a small amount of precipitate that was formed during incubation, the clear supernatant fluid was analyzed for increase in absorbance at 233nm using all the three tested strains. The average amount of reducing sugars were 0.16% of RAG-1 emulsan, 0.15% for Pay-4 emulsan and 0.14% for IL-1 emulsan, compared to glucose amine solution 1% as standard respectively (Table 5), for 15min followed by neutralization. Emulsifications were the petroleum based crude oils, while vegetable-based oils such as soya oil and microscope immersion oil were less effectively emulsified. When aromatic substrates were assayed, the emulsifying activity appeared to differ depending on the number of rings (Horacio et al., 2003). Emulsan dry mass The dry mass of emulsan made by all strains were measured. The highest amount belonged to RAG-1 which was equal to 1.89g/500mL. This amount could be considered as good result compared with other bacteria in last works (Table 6). DISCUSSION Production of emulsan depends on several factors specially carbon sources. Emulsification activity usually depends on miscelle production. In our study, it was found that the highest emulsification activity is 100% when supernatant of produced emulsan is able to form small micelles in all parts of crude oil. These results are being seen in 1% soya oil medium while in some previous work the value of 100% had been seen in 2% ethanol and 3% crude oil media (Nemati and Mazaheri Assadi, 2002). In the previous study, we used 1g/L of corn oil according to the methods of Lindhardt et al., 1998 was used. This is very much compatible with the results found by (Rubinovitz et al., 1982). The biosurfactant biosynthesis from crude oil microorganisms occurred predominantly during the exponential growth phase. Among these compounds soya oil was the best, yielding 3.4g/L the cell growth in soya oil media (Table 1). Other low molecular weight carbon sources might also be employed, but their efficacy would have to be established on a strain-by-strain basis. The results from Table 1, is enough to prove and show the cell dry weights of each strain grown on different carbon sources after 72h. These results confirm the previous work of (Amirian et al., 2004), suggesting that the biosurfactant is produced as a primary metabolite accompanying cellular, biomass formation. Furthermore, as results showed the soya oil medium has the highest emulsification activity. These results indicate that the bioemulsan produced by the isolates had high emulsification activity. These results are in accordance with the results of a substantially cell-free medium from a bacterial cell culture in which Acinetobacter calcoaceticus NRRL B-15850 has been grown, characterized by no dialyzable emulsifying activity for at least one hydrocarbon. Findings suggested that the emulsifiers activity depends on its affinity for hydrocarbon substrates which involves a direct interaction with the hydrocarbon itself rather than an effect on the surface tension of the medium (Amirian, 2004). Results indicated that the biosurfactant produced by the isolates had high emulsification specificity toward soya oil. The diameter of hemolytic zone was measured after 24h. The result showed that, the hemolytic zone for Soya was more than crude oil and ethanol had the fewest diameters. Soya oil medium showed the largest hemolytic zone while in some pervious work crude oil hsd showed the lagest zone. The obtained partially purified bioemulsifer from cell free supernatant and crude medium of all the three used strains revealed that cells grown on soya oil had the highest protein content as compared to crude oil and ethanol medium. Aqueous solutions in sea water or fresh water (the latter containing a suitable divalent cation, such as magnesium) of emulsans are excellent emulsifying agents. The results of this study showed that these three bacteria have the same ability to make an emulsan as a good emulsifier. A fraction of cell-free protein was also demonstrated to be associated with the emulsan; this is in agreement with the results found by (Shabtai and Gutnick, 1985), who checked the cell free esterase. Thus, the protein portion plays a crucial role in the emulsifying activity of the emulsan. The protein mixture associated with emulsan contains inhibitory proteins which could affect the activity towards many of substrates. Emulsifications were the petroleum-based crude oils, while vegetable-based oils such as soya oil and microscope immersion oil were less effectively emulsified. When aromatic substrates were assayed, the emulsifying activity appeared to differ depending on the number of rings (Horacio et al., 2003). Preliminary evidence suggests that crude emulsan preparations contain such proteins, and efforts to isolate and characterize them are currently in progress. In this study two isolates were screened with a potential of the highest biosurfactant production. These strains produced a bioemulsan like biosurfactant. Acientobacter calcoaceticus cells accumulated capsular material on the cell surface during logarithmic phase and then released this polymeric material in the form of an active emulsifier in stationary phase or during conditions of unbalanced growth. The result of this study is consistent with the results of Kaplan and Rosenberg., 1982. Although, bioemulsifier production by microorganisms is generally associated with growth on hydrocarbons (Rubinovitz et al., 1982), A. calcoaceticus RAG-1 (standard strain) produces at least much emulsan when grown on soya oil and Crude oil rather than on ethanol medium. Maximum biomass concentration was achieved after 32-36h of growth. Growth rate 0.24/h and maximum biomass obtained were much higher for this culture than for other biosurfactant-producing microorganisms on hydrocarbons reported in the literature (Rosenberg, 1979). Results of emulsification activity indicated that produced biosurfactant by two isolates had a high emulsification specificity with soya oil and this result differd to that of (Kaplan and Rosenberg, 1982). These findings suggest that the emulsifier's activity depends on its affinity for hydrocarbon substrates which involves a direct interaction with the hydrocarbon itself rather than an effect on the surface tension of the medium. Both types of the biosurfactants remained stable after exposure to high temperatures of 100oC and 120oC for 15min (Kim et al., 2000). All strains were tested for hemolytic activity, which is regarded by some authors as indicative of biosurfactant production and used as a rapid method for bacterial screening (Tabatabaee et al., 2005). After hemolytic test, stabilization of an oil and water emulsion is commonly used as a surface activity indicator. Several studies are focused on high emulsifying abilities Francy et al., 1991). In this study two previously isolated, high surfactant produced bacteria (i.e. PAY-4 and IL-1) were screened and compared with Acinetobacter calcoaceticus RAG1. These strains produced a bioemulsan accumulated capsular material on the cell surface during logarithmic phase and then released this polymeric material in the form of an active emulsifier in stationary phase or during conditions of unbalanced growth similar to the findings of (Kaplan et al., 1982). Although bioemulsifier production by microorganisms is generally associated with growth on hydrocarbons, Acinetobacter Calcoaceticus RAG-1 (standard strain), PAY-4 and IL-1 produces more emulsan when grown on soya oil and crude oil rather than on n-hexadecane and ethanol medium (Table 6). These result are similar to results obtained by (Shabtai, 2005).Many other studies are needed to complete the structural analysis Fig. 2 of such macromolecules. ACKNOWLEDGEMENTS The help rendered by Mr. Sheikhi Nejad at Biotechnology Department of Iranian Research Organization for Sciences and Technology is highly appreciated. REFERENCES
© 2008 Tehran University of Medical Sciences Publications The following images related to this document are available:Photo images[se08020t4.jpg] [se08020t6.jpg] [se08020f2.jpg] [se08020t3.jpg] [se08020t5.jpg] [se08020t1.jpg] [se08020f1.jpg] [se08020t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}