 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
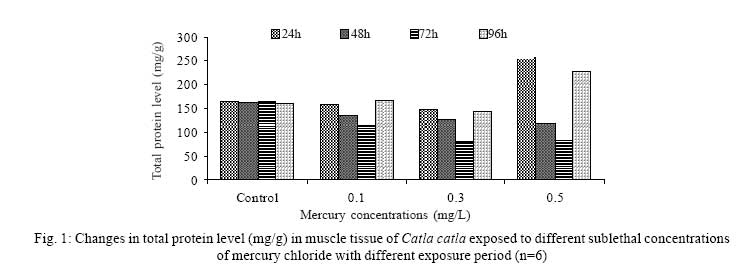
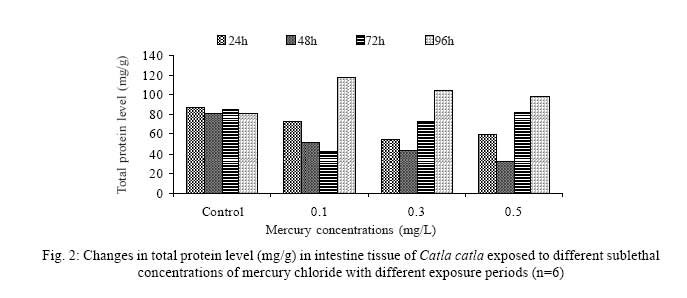
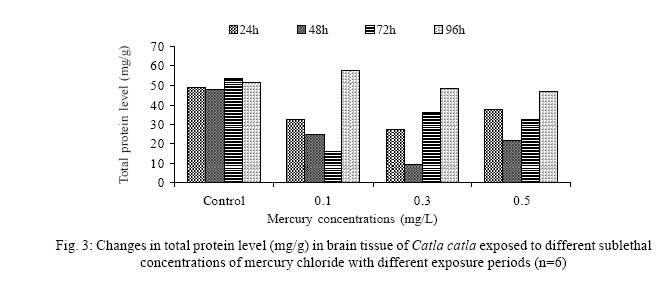
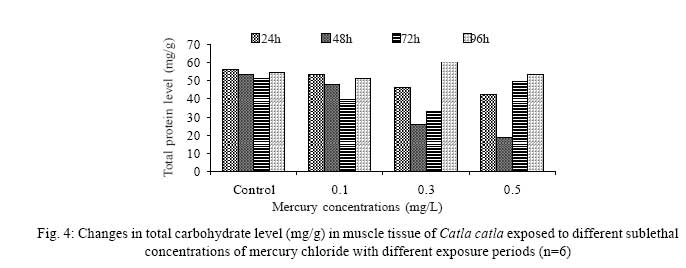
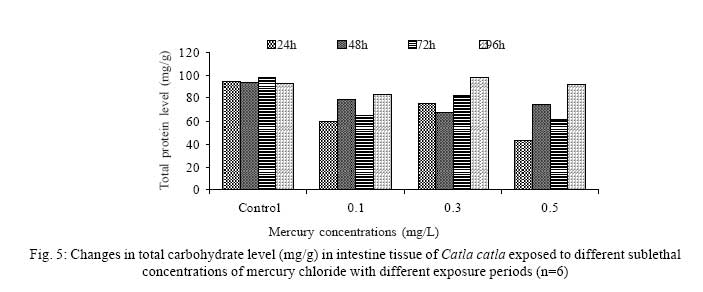
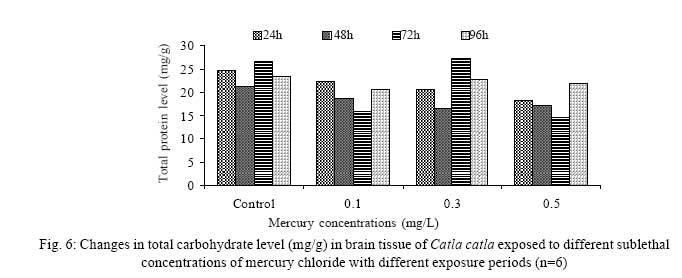
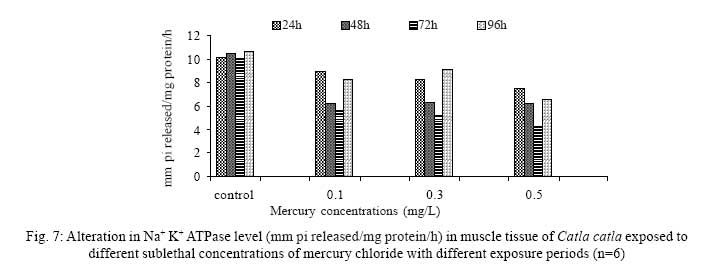
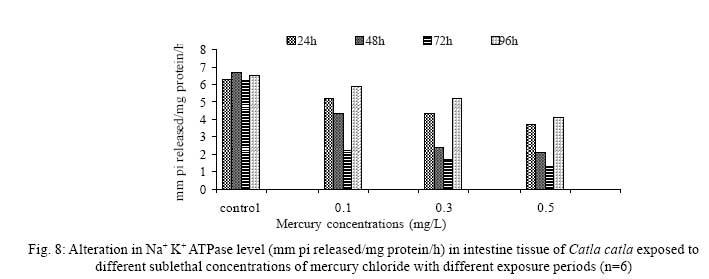
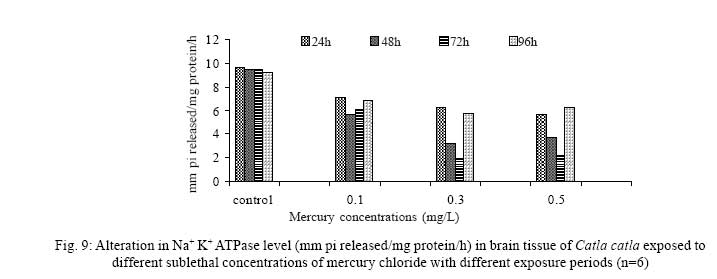
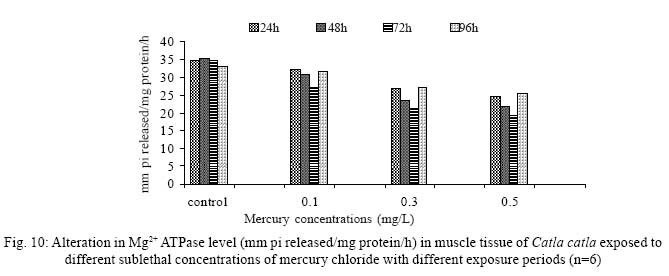
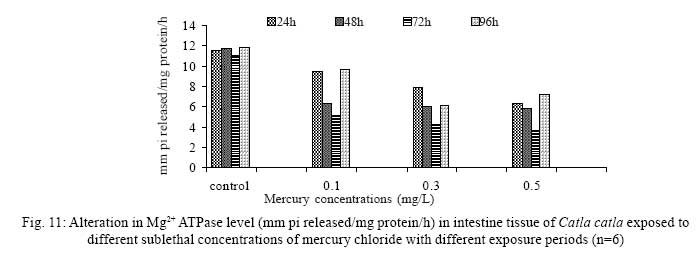
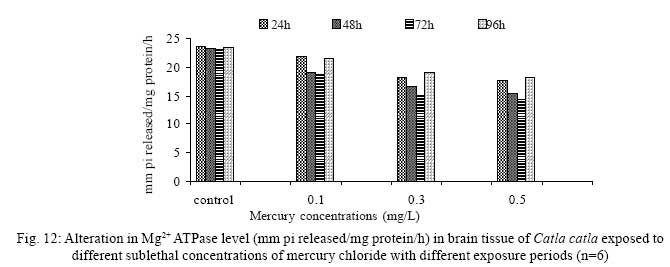
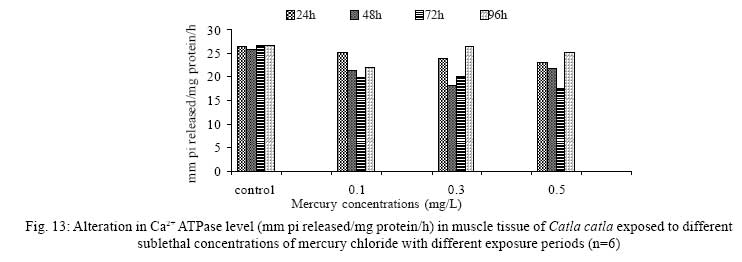
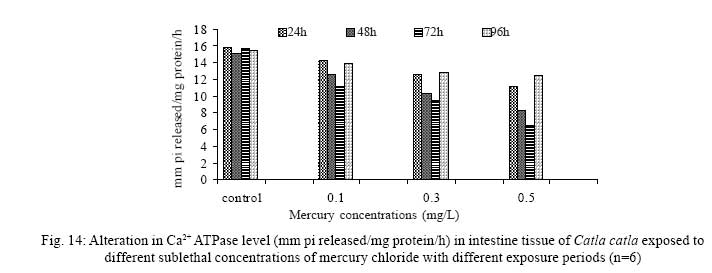
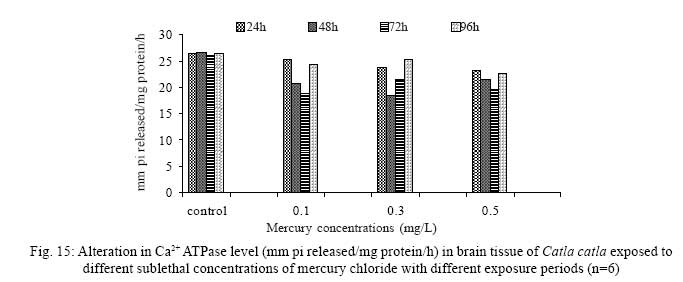
Iranian Journal of Environmental Health Science & Engineering, 2008, Vol. 5, No. 2, pp. 109-116 Biochemical Study of Freshwater Fish Catla catla with Reference to Mercury Chloride 1 P. Martin Deva Prasath, *2 S. Arivoli 1 Department of Chemistry, TBML College, Porayar-609 307, Nagai District, Tamil Nadu, India Received 16 March 2007; revised 18 September 2007; accepted 15 February 2008 Code Number: se08021 ABSTRACT Toxic activity of mercury chloride was tested in vivo on fresh water fish Catla catla. Acute toxicity testswere conducted to measure the impact of toxicity on the fishes within a short period at the variousconcentrations of HgCl2 (0.1, 0.5, 1, 1.5, 2, 2.5 and 3mg/L). The protein and carbohydrate were estimatedusing Anthrone by standard methods and enzymes such as Na+-K+ , Mg2+ and Ca2+ adenoxidetriphosphatases were determined caloriemetrically. Depletion of protein was observed at all exposureperiods. The biochemical estimation values of carbohydrates in muscle, intestine and brain showedsignificant values with P<0.05. The activity of Mg 2+ adenoxide triphosphatases in muscle, intestine andbrain was studied and the maximum depletion was observed at 96h, 72h and 72h, respectively. Regardingactivity of Ca 2+ adenoxide triphosphatases in muscle, the maximum depletion of this enzyme activity inmuscle was observed at all exposure periods, except 96h. In intestine, enzyme activity was graduallydecreased in all exposure and an increase was found during 96h. In brain, the enzyme activity wasobserved at all exposure periods except 96h. Key words: Toxic, protein, carbohydrate, enzyme, Catla catla INTRODUCTION Heavy metals are being passed on into aqueous environments through industrial processes, sewage disposal, soil leaching and rainfall. The concentrations of these heavy metals are sub- lethal or lethal to aquatic organisms when the duration of exposure to these metals are prolonged (Eisler and Gardener, 1973). It is well documented that effect of heavy metals are dependent upon the physical and chemical conditions of the environment especially water salinity, hardness, pH and dissolved oxygen and can act synergistically. Heavy metals from several industrial mining and other sources enormously contribute to the pollution problem in rivers and streams resulting in adverse impacts on biota including fish. Fish population is generally considered very sensitive to all kinds of environmental changes to which it is exposed as they are exclusively aquatic with external mode of fertilization. Certain stages in the life cycle of fresh water fish are more susceptible to environmental and pollution stresses (Von Westernhagan, 1988). Even though many studies are available in toxicity of various fishes (Jagadeesan et al., 1991; Margarat et al., 1991; Jagadeesan et al., 2001; Francis et al., 2002;), there is no report available on Catla catla. Hence the present study has been carried out on Catla catla with reference to mercury chloride. MATERIALS AND METHODS Finger lings of Catla catla species were collected and kept in aquarium and the water used was clear and unchlorinated (APHA, 1980). Fishes were fed daily with fish pellets and acclimatized for 30 days. Acute toxicity tests were conducted to measure the impact of toxicity on the fishes within a short period (Sprague, 1969) at the various concentration of HgCl2 (0.1, 0.5, 1, 1.5, 2, 2.5 and 3mg/L). The tissue was homogenized with 1mL of Tris-HCl buffer. The protein and carbohydrate were estimated by standard methods (Carroll et al., 1956) and enzymes such as Na+-K+, Mg2+ and Ca2+ adenoxide triphosphatuses were analyzed by caloriemetric method (Fiske et al., 1925). RESULTS Protein Depletion in the protein content in the muscle, intestine and brain of the (Catla catla) exposed to mercury chloride for 24h, 48h, 72h and 96h in 0.1, 0.3 and 0.5m/L sub-lethal concentrations were estimated. Protein content in the muscle of the control, depletion and increase of muscle protein are shown in Fig. 1. Intestine of the mercury chloride treated fish shows a gradually decrease in protein level. Depletion of protein was observed at all exposure periods (Fig. 2). The level of protein in the control fish brain, is presented in Fig. 3. Carbohydrates Depletion of carbohydrate content of the muscle (Fig. 4), intestine (Fig. 5) and brain (Fig. 6) of catla catla exposed to the mercury chloride for 24, 48, 72 and 96h in 0.1, 0.3 and 0.5mg/L sublethal concentrations were estimated. Among these, the maximum depletion of carbohydrate was observed in intestine during 96h. Generally, depletion in carbohydrate content is directly proportional to the exposure period of the toxicant. The obtained biochemical estimation values of the muscle, intestine and brain were subjected to statistical analysis and showed significant values at P<0.05. Enzyme analysis The activity of Na+-K+, Ca2+ and Mg2+ ATPase in muscle (Fig. 7), intestine (Fig. 8) and brain (Fig. 9) were estimated in the experimental fish after 24, 48, 72 and 96h in 0.1, 0.3 and 0.5mg/L sub-lethal concentration of exposure to the mercury chloride. The alteration in the activity of enzymes of Na+ K+ ATPase were observed. The activity of Mg2+ ATPase in muscle (Fig. 10), intestine (Fig. 11) and brain (Fig. 12) were studied and the maximum depletion was observed in 96h, 72h, and 72h, respectively. The activity of Ca2+ ATPase in muscle and depletion of enzyme activity was observed at all exposure periods, except for 96h (Fig. 13). In intestine, enzyme activity was gradually decreased in all exposures and increased during 96h (Fig. 14). In brain, the enzyme activity was also observed at all exposure periods except 96h (Fig. 15). DISCUSSION Biochemical AnalysisProtein Proteins are involved in major physiological events therefore the assessment of the protein content can be considered as a diagnostic tool to determine the physiological phases of organism. Proteins are highly sensitive to heavy metal poisoning (Jacobs et al., 1977). Depletion of protein content has been observed in the muscle, intestine and brain of the fish Catla catla as a result of mercury chloride toxicity. When an animal is under toxic stress, diversification of energy occurs to accomplish the impending energy demands and hence the protein level is depleted (Neff, 1985). The depletion of total protein content may be due to breakdown of protein into free amino acid under the effect of mercury chloride at the lower exposure period (Shakoori et al., 1994). Corresponding to it a level of increase in protein content was observed at increased exposure period of 96h at 0.1, 0.3 and 0.5mg/L sublethal concentration. These indicate that mercury induces proteolysis in the fish even under sublethal toxic stress resulting in elevated levels of protein content; but, the degree of proteolysis appears time-dependent, as the decrease in protein levels progressed significantly at 24h, 48h and 72h but regressed and attained almost normally at 96h of exposure. Metals could alter the structure, permeability and integrity of lysosomal membranes resulting in the diffusion of their enzyme into cytosol (Sternlib et al., 1976). Hence high activity of protease, a lysosomal enzyme, in the organs of fish might be due to the damage caused by mercury to lysosomes. Elevated protease activity induced proteolysis, the intensity increased with the increase in exposure period from 25h to 72h may be the increase in free amino acid pool due to increased proteolysis would act as an osmotic and ionic effectors to bring the electrostatic equilibrium between the external medium and blood (Schmit-Nielson, 1975; Jurss, 1980). Besides, free amino acids would also serve as precursors for energy production under stress, and for the synthesis of required proteins to face the metal challenge (Sree devi et al., 1991). Carbohydrate In the present study, an initial decrease and an increase in the level of total carbohydrate has been noticed in the muscle and intestine tissues. The disturbance in the carbohydrate metabolism was considered as one of the most outstanding biological lesions due to the action of heavy metal (De Bruin, 1976). The decrease in carbohydrate content in the muscle, intestine and brain may be due to glucose utilization to meet excess energy demand imposed by severe anaerobic stress of mercury intoxication (Margarat et al., 1999). Another possible reason for depletion in the tissue may be due to impairment of glycogen synthesis. Under hypoxic conditions; fish derive the energy by anaerobic breakdown of glucose which is available to the cells with the increased glycogenolysis. The observed depletion of carbohydrate in the present study explains the increased demand of these molecules to provide energy for the cellular biochemical process under toxic manifestations. Similar results were observed in Thalmile crenata, Anabas testudienues and Anabas scandens, when exposed to copper, lead, nitrate and mercury chloride, respectively (Villalan et al., 1988; Mary Candravathy et al., 1991). During the initial period, metabolic activity failed to recover indicating the effects of accumulated mercury in the tissues. However, at 96h the metabolites reached to normal levels. It indicates the slow elimination of mercury and resynthesis of metabolities against the toxicants, indicating the reduced rates of glycogenolysis and glycolysis. The recovery could be attributed to restoration of regulatory function of phosporylases by elimination of toxicant (Holcombe et al., 1976; Richert et al., 1979) from the endocrine glands like pancreas and adrenals. Enzyme analysisAdenosine triphosphatases (ATPases) The major target molecules affected by metals include ion dependent ATPases, which lead to disturbances in ion homeostasis. The inhibition of ATPases lead to decreased ATP breakdown and reduced the availability of free energy. The reduced energy supply may affect several metabolic processes (Ramalingam et al., 1999). Na+K+-ATPase Na+K+-ATPase is considered as a marker enzyme to understand the physiological impairment of the cell (Campbell et al., 1974). The inhibition of membrane bound ATPases in the tissues, muscle, intestine and brain could be result of physico-chemical alteration of the membrane. The alteration in ionic balance depolarises the nerve and due to depolarisation the nerve cells increase in releasing the neuro transmitter which inturn inhibits Na+K+-ATPase activity (Kimellberg et al., 1974). Inhibition of the enzyme increases intracellular Ca2+ and decreases intracellular Mg2+ concentration (Karamer et al., 1991). Ca2+ ATPases Calcium ions are essential for the transmitter release and the level of calcium is regulation by Ca2+ ATPase (Rubin, 1970). Ca2+ ATPase maintains low intracellular Ca2+ ions than the Ca2+ concentration of extra cellular medium (Harrison et al., 1980). Decreased Ca2+ ATPase due to the activity of Mercury chloride may lead to high internal Ca2+ level (Duncan, 1967). Increase in intracellular accumulation of Ca2+ ions there by increases the release of neuro-transmitter from the synaptic vesicles by exocytosis (Yamaguchi et al., 1979) which may inhibit Na+K+ ATPase. Mg2+ ATPases Mg2+ ATPase is involved in the control of passive permeability (Duncan, 1967). It is also involved in oxidative phosphorylation, the inhibition due to toxicants directly prevents or reduces the oxidative phosphorylation (Nuefeld et al., 1979). Inhibition of Mg2+ ATPase may result in effects on energy metabolism and respiration (Desaiah et al., 1977). During the initial exposure period, metabolities activity failed to recover indicating effects of accumulated mercury in the tissues. However on the 96 hr of exposure, the metabolities reached to near normal levels. Generally, more energy is needed to mitigate any stress condition and this may be obtained from carbohydrates and proteins. Increase in structural proteins could help the animal to fortify its organs for developing resistance and increase in soluble fraction from the general intracellular environment and help the animal to adapt to the imposed toxic stress Sivaramakrishnan et al., 1998). ACKNOWLEDGEMENT The authors thank the Heads of Department of Zoology and chemistry for their encouragement. REFERENCES
© 2008 Tehran University of Medical Sciences Publications The following images related to this document are available:Photo images[se08021f9.jpg] [se08021f3.jpg] [se08021f12.jpg] [se08021f13.jpg] [se08021f8.jpg] [se08021f11.jpg] [se08021f4.jpg] [se08021f2.jpg] [se08021f1.jpg] [se08021f15.jpg] [se08021f7.jpg] [se08021f6.jpg] [se08021f5.jpg] [se08021f10.jpg] [se08021f14.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}