 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Iranian Journal of Environmental Health, Science and Engineering, 2009, Vol. 6, No. 1, pp. 17-22 Chromium Induced Cytotoxicity in Blackgram (Vigna Mungo L.) A. Chidambaram, P. Sundaramoorthy, A. Murugan K. Sankar Ganesh, *L. Baskaran
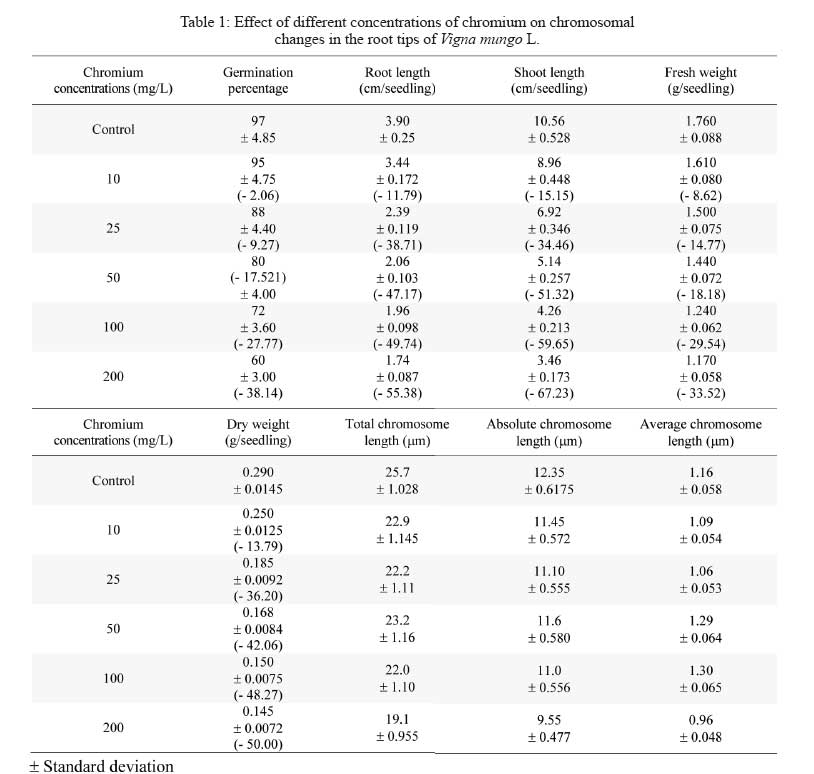
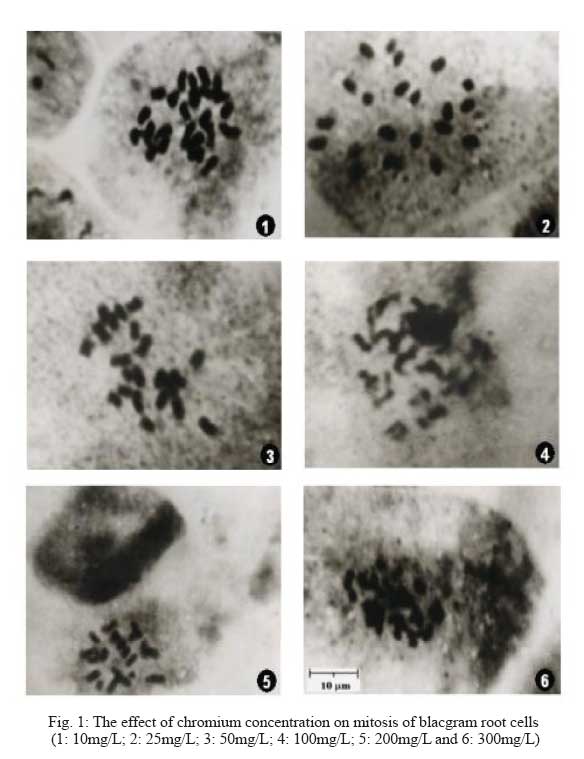
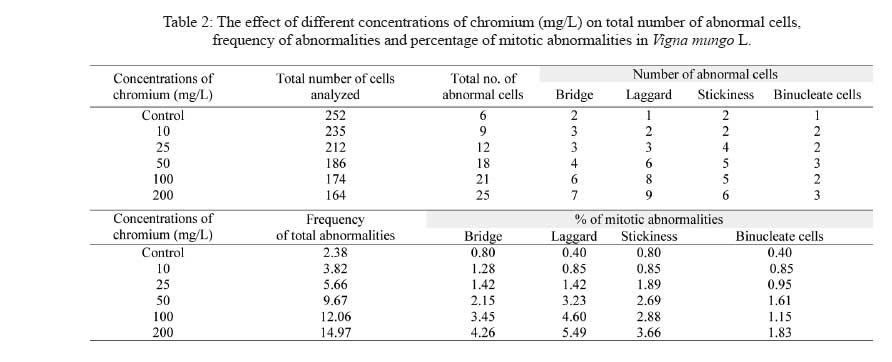
Division of Environmental Biology, Department of Botany, Annamalai University, Annamalai Nagar 608002, Tamil Nadu, India Code Number: se09004 Received 17 September 2007; revised 18 August 2008; accepted 24 September 2008 ABSTRACT Chromium is known to be highly toxic to biological systems. This study was designed to determine the mutagenic effects of different concentrations (0, 10, 25, 50, 100 and 200 mg/L) of hexavalent chromium on root tip cells of blackgram (Vigna mungo L. Hepper). The blackgram seeds were equi-spacially arranged in sterilized petriplates lined with filter paper and they were treated with different concentrations of chromium solution. In germination studies, the morphological growth parameters such as germination percentage, root length, shoot length fresh weight and dry weight of blackgram seedlings were decreased with increasing dose of chromium concentrations. No germination of blackgram seeds was recorded at 300mg/l chromium concentration. Chromosome aberration assay was used to determine the mitotic indices and rate of chromosome aberration in blackgram root tip cells due to chromium treatment. The results showed that the mitotic indices were complicated due to different concentrations of chromium. However, the increase in chromium concentration has led to a gradual increase in the percentage of chromosomal aberration and mitotic index. The chromosome length, absolute chromosome length and average chromosome lengths were gradually found to decrease. There was no considerable change in 2n number of chromosome with the increase in chromium concentrations. It is concluded that the hexavalent chromium has significant mutagenic effect on the root tip cells of blackgram. Key words: Hexavalent chromium, cytotoxicity, blackgram, chromosomal aberration, germination, mutation INTRODUCTION Heavy metal pollution of water is a major environmental problem facing the modern world. The global heavy metal pollution is increasing in the environment due to increase in number of industries. Many industrial wastewaters contain heavy metals like cadmium, lead, zinc, cobalt and chromium. Among heavy metals, chromium plays a major role in polluting our water environment. Chromium can co-exist in the environment in two oxidation states viz., trivalent chromium and hexavalent chromium. The hexavalent chromium is released from various industries such as electroplating, leather tanning, textile printing, textile preservation and metal finishing. The compounds of chromium have been known to be strong carcinogens and mutagens that can reach the target organs of human through drinking water. Chromium is often admixed with industrial effluents that are used for irrigation. The uptake of excess concentrations of heavy metals, reduced the growth of plants (Abdel-Azeem and El-Nahas, 1996; Salem, 2000). The effluent irrigation adversely affect plant growth and development (Shanker et al., 2005, Yongpisanphop et al., 2005, Panda and Choudhury, 2005). This alternation in plant growth is correlated with the disruption of the physiological and cytological processes in plant cells. By this way the processes of respiration, photosynthesis and mitotic activity are greatly affected by the toxic effect of heavy metals (Ewais, 1997). Previous studies showed that chromium in higher concentrations severely affected the germination, growth and yield of plants (Lakshmi and Sundaramoorthy, 2003; Purohit et al., 2003; Sankar Ganesh et al., 2006; Chidambaram et al., 2006; Sundaramoorthy and Sankar Ganesh, 2007). The research work on the effect of hexavalent chromium on chromosomal behaviour in plant cells are very rare so this research attempt has been made to deal with the effect of different concentrations of hexavalent chromium on the mitotic index and the aberration rate of in chromosome blackgram (Vigna mungo L.) root tip cells. MATERIALS AND METHODS The seeds of blackgram (Vigna mungo L. Hepper) seeds were collected from Tamil Nadu Rice Research Institute, Regional Research station of Tamilnadu Agricultural University, Aduthurai, Tanjore district of Tamil Nadu. Chromium stock solution was prepared by dissolving 141.4 mg of potassium dichromate (K2Cr2O7) in 100 mL distilled water. From the potassium dichromate stock solution, then different concentrations of 5, 10, 25, 50, 100, 200 and 300 mg/L of chromium solution were prepared freshly at the time of experiments. Uniform size of healthy blackgram seeds were equi-spacially arranged in sterilized petriplates lined with filter paper. The seeds were treated with equal volumes (20 mL) of different concentrations (10, 25, 50, 100, 200 and 300mg/L) of chromium solution. One set of seeds irrigated with distilled water was maintained as the control. The emergence of radicle was noticed after two days and they were used for cytological studies. The root tips were collected from the both control and treated seedlings were than washed in distilled water and fixed in 1:3 acetic alcohol. The root tip squashes were made by using, haematoxylin squash technique of Marimuthu and Subramanian (1960). The root tips were hydrolysed in 0.1 N HCl for 15 to 20 min at 60°C and then they were thoroughly washed in distilled water and treated with 4% iron alum (Ferric ammonium sulphate) for 5 min. The root tips were then washed thoroughly in distilled water and transferred in dilute haematoxylin stain and kept for three hours. The root tips were thoroughly washed in distilled water and then they were treated in 45% acetic acid for a few seconds to soften the tissues. One or two root tips were placed on the tissues clean slide and squashed by using a cover slip and the slide was sealed and mounted. The slides were observed under microscope to find out the structural changes in chromosomes due to chromium treatment. The variation between the control and the treated total chromosomal length, absolute and average chromosomal length were noted. Similarly, the percentage of mitotic abnormalities were also observed in treated plants. The chromosomal aberrations were examined and they were counted and microphotographed from the squashes. Each treatment was repeated at least three times as described above. RESULTS The gradual decrease of germination percentage, root length, shoot length, fresh weight, dry weight, total chromosome length, absolute chromosome length and average chromosome length of blackgram seedlings with increase of chromium concentrations are given in Table 1. Higher seed germination (97.0%), root length (3.90 cm/seedling), shoot length (10.56 cm/seedling), fresh weight (1.760 g/seedling), dry weight (0.290 g/seedling), total chromosome length (25.7 µm), absolute chromosome length (12.35 µm) and average chromosome length (1.16 µm) were observed in the root tips of control seedlings. Similarly, the lower germination percentage (60.0 %), root length, (1.74 cm/seedling) shoot length (3.46 cm/seedling), fresh weight (1.170 g/seedlings), dry weight (0.145 g/seedlings) total chromosome length (19.2 µm), absolute chromosome length (9.55 µm) and average chromosome length (0.94 µm) were observed at 200 mg/L chromium concentrations. The reduction of chromosome length was observed at 10 to 200 mg/L chromium concentrations onwards. Similar findings were observed in earlier reports (Vasquez et al., 1986; Salem et al., 1993). The frequency of total abnormalities (2.38, 3.82, 5.66, 9.67, 12.06 and 14.97) was observed in various concentrations (control, 10, 25, 50, 100 and 200 mg/L) of chromium, respectively. The number of abnormal cells having bridge, laggard, stickiness and Binucleate cells are gradually increased from control to 200 mg/L onwards. The similar findings were observed by Levis and Bianchi (1982). Analysis of various stages of cell division revealed that reduction of 93.08% in metaphase and 90.55% in anaphase stage at 80 ppm as compared to 26.76% decrease of metaphase and 7.3% anaphase at lower dose of chromium of blackgram seedlings. This effect depends upon the concentration given to the seeds. Heavy metals (Cd and Pb) are extremely toxic elements and they could reduce the mitotic activity and induce many types of chromosomal anomalies (Mukherji et al., 1990; Gupta and Gupta, 1992; Abu Saidah, 1995; Abdel-Azeem and El-Nahas, 1996). The increasing concentrations of the heavy metal induce a marked reduction in the mitotic activity of root meristems Vicia faba where as the total percentage of chromosomal aberrations were found to be increased. Chromosomal aberrations in meristermatic cells of roots show stickiness in chromosomes and disturbances that C-metaphase may affect the spindle apparatus (Grant, 1978). Chromosomal stickiness could also be observed at high frequency in this study. The induction of this type of aberration may be due to the disturbance in nucleic acid metabolism in the cell. The reduction of chromosome length was observed at 10 mg/L to 200 mg/L chromium concentrations onwards. Similar findings were observed in earlier reports (Vasquez et al., 1986, Salam et al., 1993). The growth inhibition at higher concentrations may be linked with lower mitotic activity in the root meristematic zone. It may also be due to an inhibition of cell enlargement in the elongation zone under heavy metal stress (Gabbrielli et al., 1990). The total number of abnormal cells (bridge, laggard, stickiness and binucleate cells), frequency of total abnormalities and percentage of mitotic abnormalities gradually increased with the increase of chromium concentrations (Table 1 and Fig. 1). But, the total number of cells gradually decreased with the increase of chromium concentrations. The total number of cells (252, 235, 212 186, 174 and 164) were observed in the root tips grown in the various concentrations (10, 25, 50, 100 and 200 mg/L) of chromium respectively. The total number of cells (252, 235, 212 186, 174 and 164) were observed in the root tips grown in the various concentrations (10, 25, 50, 100 and 200 mg/L) of chromium, respectively. It may be attributed to enhance disturbance of spindle function with increase in chromium conentration (Jain et al., 2000). The hexavalent chromium is s highly bioactive compound can easily enter the cell through cell membrane and it generates some active oxides. These active oxides can combine with the intracellular DNA and lead to the unreliable intercrossing connection and duplication in DNA and ultimately result in chromosomal aberration (Li et al., 1995; Qian, 2004). Thus, it has cytotoxic effects which can even lead to DNA damage (Edwards and Karen, 1994). The number of abnormal cells such as bridge, laggard, stickiness and Binucleate cells are gradually increased from control to 200 mg/L onwards. The similar findings were observed in Allium cepa L. (Jayaprakash et al. 1994) and sugarcane (Jain et al. 2000) due to chromium treatment. Analysis of various stages of cell division revealed that reduction of 93.08% in metaphase and 90.55% in anaphase stage at 80 ppm as compared to 26.76% decrease of metaphase and 7.3% anaphase at lower dose of chromium (1 ppm). The high doses of chromium supply has a toxic effect on cell division attributes. The increased number of chromosomal aberrations like stickiness, laggards, chromosome bridges, C-metaphase, fragmentation and binucleate cells were observed with the increasing concentrations of metals (Table 2). Fig. 1 shows the effect of chromium on mitosis of blacgram root cells. DISCUSSION The germination percentage, root length, shoot length, fresh weight, and dry weight of blackgram gradually decreased with the increase in chromium concentrations. This may be due to the accumulation of chromium in seeds (Sankar Ganesh et al., 2006). In the present study, the different concentrations of hexavalent chromium could lead to various forms of chromosomal aberrations. The aberration rate depends upon the chromium concentrations. The chromosomal aberration might be induced directly by chemical compounds. It may also be induced by disturbing the process of DNA and protein synthesis or during the RNA translocation. It may also be induced by the activities of chemical compounds to prevent the re-establishment of the chromosome under normal repairing of some damage. The occurrence of many chromosomal aberrations in the experiment clearly indicate that the hexavalent chromium has a genotoxical effect on root cells of blackgram seedlings. This effect depends upon the concentration given to the seeds. Heavy metals (Cd and Pb) are extremely toxic elements and they could reduce the mitotic activity and induce many types of chromosomal anomalies (Mukherji et al., 1990; Gupta and Gupta, 1992; Abu Saidah, 1995; Abdel-Azeem and El-Nahas, 1996). The increasing concentrations of the heavy metal induce a marked reduction in the mitotic activity of root meristems Vicia faba where as the total percentage of chromosomal aberrations were found to be increased. Chromosomal aberrations in meristermatic cells of roots show stickiness in chromosomes and disturbances that C-metaphase may affect the spindle apparatus (Grant, 1978). Chromosomal stickiness could also be observed at high frequency in this study. The induction of this type of aberration may be due to the disturbance in nucleic acid metabolism in the cell. The reduction of chromosome length was observed at 10 mg/L to 200 mg/L chromium concentrations onwards. Similar findings were observed in earlier reports (Vasquez et al., 1986, Salam et al., 1993). The growth inhibition at higher concentrations may be linked with lower mitotic activity in the root meristematic zone. It may also be due to an inhibition of cell enlargement in the elongation zone under heavy metal stress (Gabbrielli et al., 1990). The total number of abnormal cells (bridge, laggard, stickiness and binucleate cells), frequency of total abnormalities and percentage of mitotic abnormalities gradually increased with the increase of chromium concentrations (Table 1 and Fig. 1). But, the total number of cells gradually decreased with the increase of chromium concentrations. The total number of cells (252, 235, 212 186, 174 and 164) were observed in the root tips grown in the various concentrations (10, 25, 50, 100 and 200 mg/L) of chromium respectively. The total number of cells (252, 235, 212 186, 174 and 164) were observed in the root tips grown in the various concentrations (10, 25, 50, 100 and 200 mg/L) of chromium, respectively. It may be attributed to enhance disturbance of spindle function with increase in chromium conentration (Jain et al., 2000). The hexavalent chromium is s highly bioactive compound can easily enter the cell through cell membrane and it generates some active oxides. These active oxides can combine with the intracellular DNA and lead to the unreliable intercrossing connection and duplication in DNA and ultimately result in chromosomal aberration (Li et al., 1995; Qian, 2004). Thus, it has cytotoxic effects which can even lead to DNA damage (Edwards and Karen, 1994). The number of abnormal cells such as bridge, laggard, stickiness and Binucleate cells are gradually increased from control to 200 mg/L onwards. The similar findings were observed in Allium cepa L. (Jayaprakash et al. 1994) and sugarcane (Jain et al. 2000) due to chromium treatment. Analysis of various stages of cell division revealed that reduction of 93.08% in metaphase and 90.55% in anaphase stage at 80 ppm as compared to 26.76% decrease of metaphase and 7.3% anaphase at lower dose of chromium (1 ppm). The high doses of chromium supply has a toxic effect on cell division attributes. Similar type of abnormalities is due to loss of microtubule of spindle fibres (Pickett-Heaps and Trimothy, 1982; Salem et al., 1993). The macronuclei observed at higher doses may originate from a lagging chromosome or from a chromosome fragment (Badr and Ibrahim, 1987). The increased number of chromosomal aberrations like stickiness, laggards, chromosome bridges, C-metaphase, fragmentation and binucleate cells were observed with the increasing concentrations of metals. It may due to the effect of hexavalent chromium on spindle apparatus. The formations of chromosomal stickiness could also be observed at high frequency owing to the disturbance in nucleic acid metabolism of the cell (Chidambaram et al. 2006) The accumulation of prophase cells indicates that Cr(VI) acts as prophase poison by exerting deleterious effects on the enzymatic activity of ribonuclease and inhibits the progressive cell division cycle (Jayaprakash et al., 1994). Chromosomal aberrations in meristematic cells of roots show that chromosome stickiness, disturbance, C-metaphase may be due to the effect on the spindle apparatus (Mansour and Kamel, 2005). These most abundant chromosomal changes and disturbed phases were noticed due to the metal treatment. It caused changes in the protein banding pattern of especially at the regions of high molecular weight (George, 1999). Similar findings were observed by Qian, (2004) while studying the root tip cells of Vicia faba treated with chromium. AKNOWLEDGEMENTS Authors thank Dr. R. Panneerselvam, Professor and Head, Department of Botany, Annamalai University for providing laboratory facilities. REFERENCES
© Copyright 2008 - Tehran University of Medical Sciences Publications The following images related to this document are available:Photo images[se09004t2.jpg] [se09004f1.jpg] [se09004t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}