 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Smithiana, Publications in Aquatic Biodiversity, Bulletin 2 June 2003, pp. 1-18 Revision of the gurnard fish subgenus Otohime (Triglidae: Pterygotrigla) W. J. Richards1, T. Yato2, and P. R. Last3 1 NOAA Fisheries, 75 Virginia Beach Drive, Miami, Florida 33149,
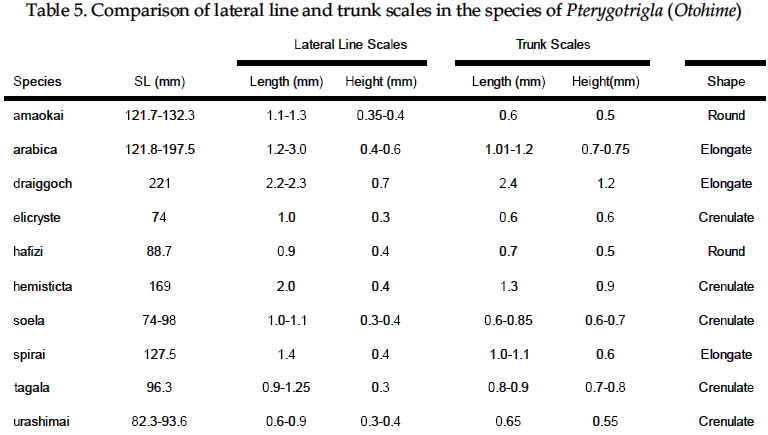
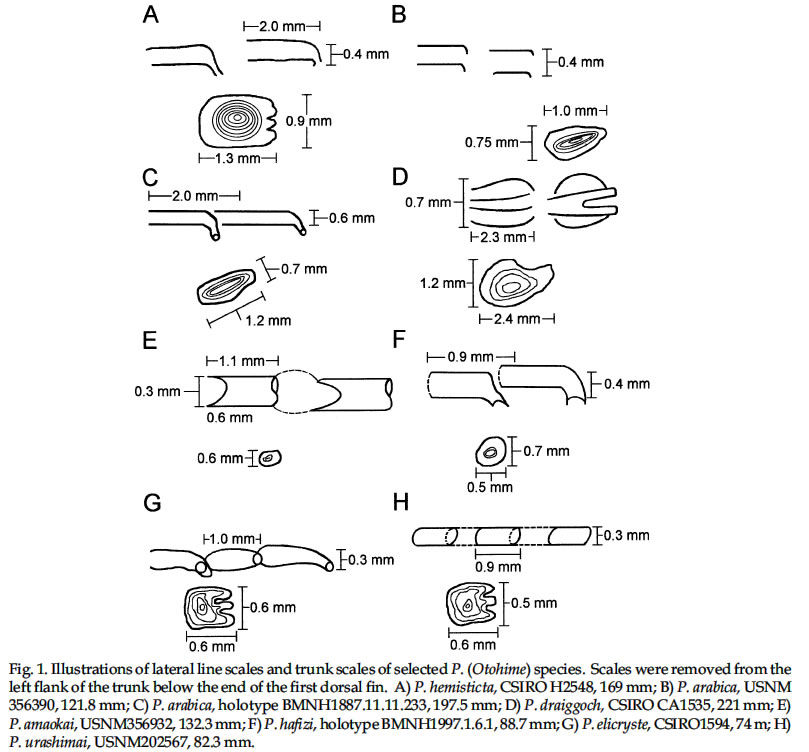
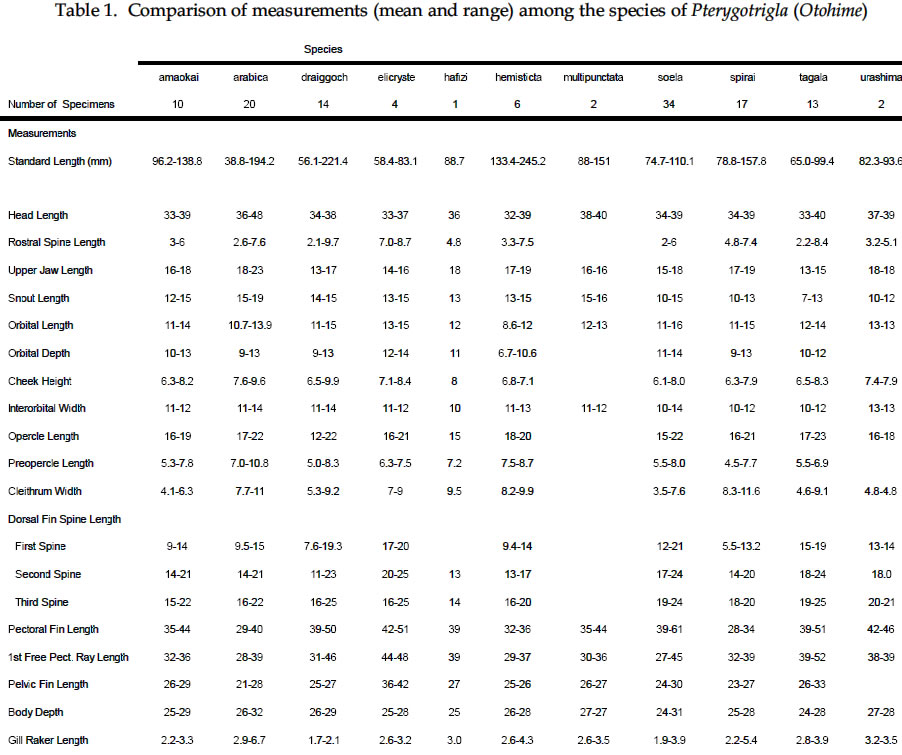
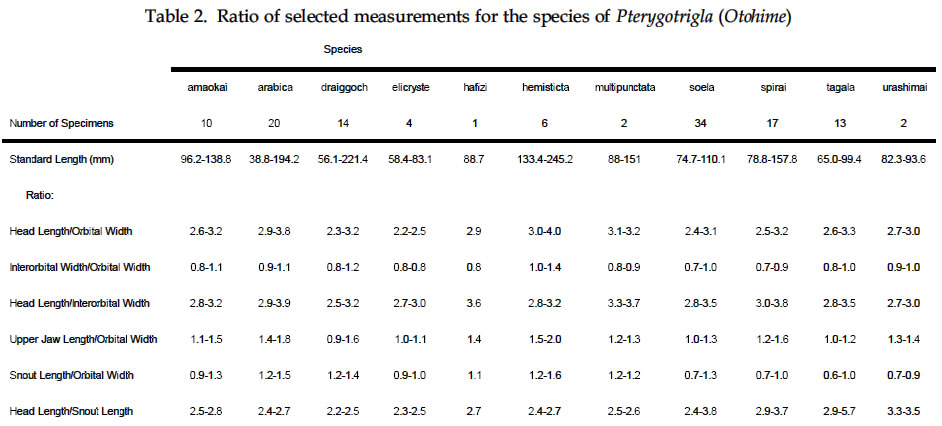
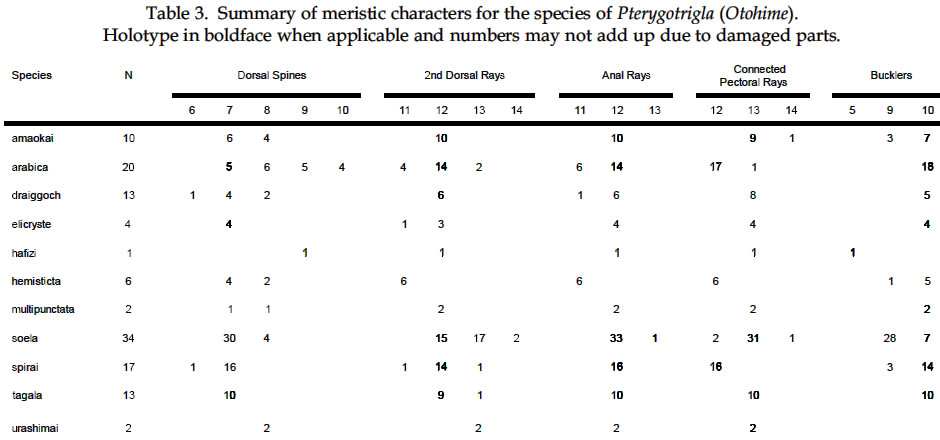
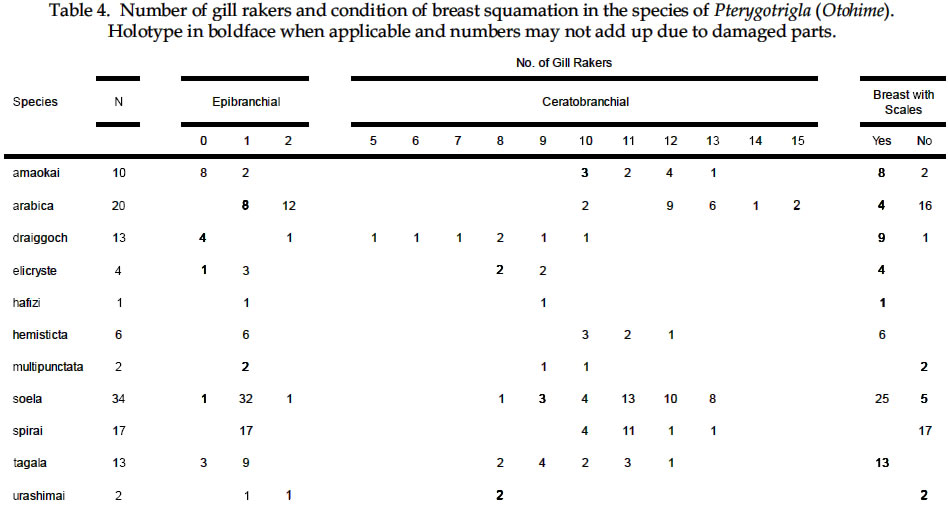
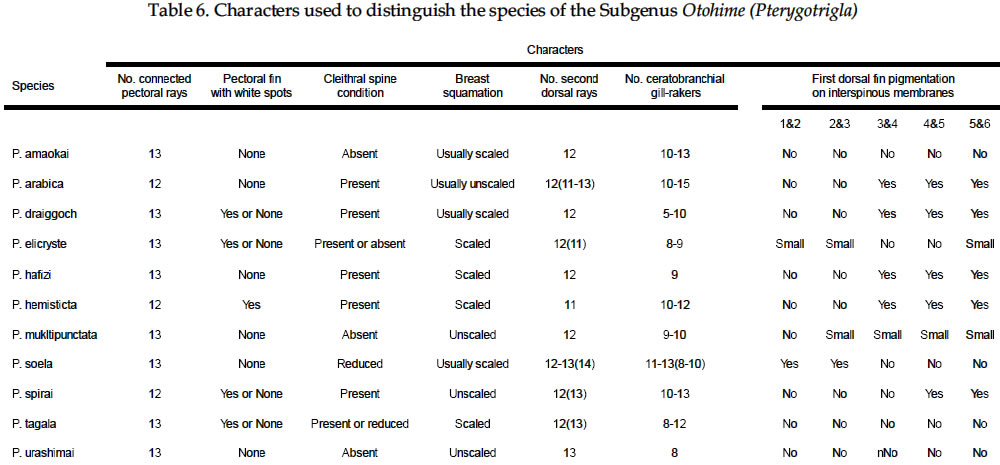
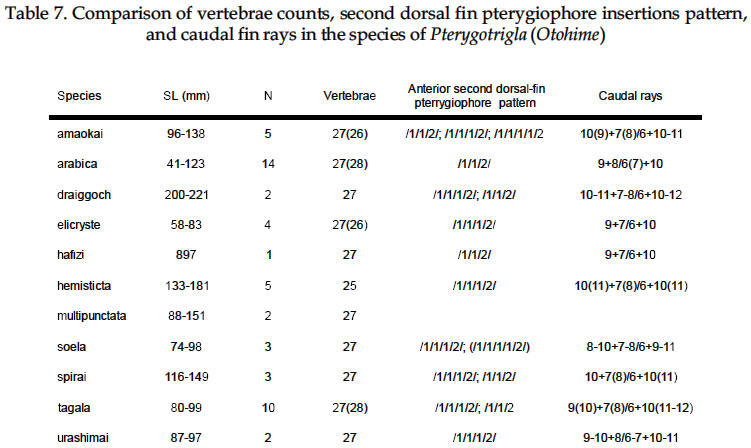
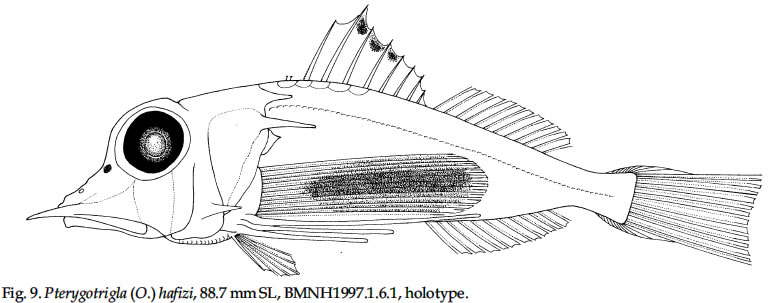
USA Code Number: sm03002 ABSTRACT Richards, William J., Takuji Yato, & Peter R. Last (2003). Revision of the gurnard fish subgenus Otohime (Triglidae: Pterygotrigla). Smithiana, Bulletin 2. The subgenus Otohime of the triglid genus Pterygotrigla is revised and includes descriptions of six new species (P. amaokai, P. draiggoch, P. elicryste, P. hafizi, P. soela, and P. urashimai) and diagnoses of five previously described species (P. arabica, P. hemisticta, P. multipunctata, P. spirai, and P.tagala). All are poorly represented in museum collections and are distributed in tropical waters of the Indian and western Pacific oceans. A brief discussion of the genus Pterygotrigla is provided together with its current species composition. The subgenus Otohime is unique within Pterygotrigla in having a very long opercular spine and cleithral spines reduced or absent . The species characters used for identification are the number of joined pectoral-fin rays and second dorsal-rays, colouration of the pectoral fin and first dorsal fin, breast squamation, and number of gill rakers and a few other meristic and morphometric features. Otohime species are very similar in morphometry and meristics and the extent of intraspecific variation is indeterminable from the small collections available. A brief discussion of the genus Pterygotrigla is provided together with its current species composition that is thought to include the unresolved triglid Prionotus alepis. A range extension is given for Pterygotrigla macrorhynchus. The Indo-Pacific triglid genus Pterygotrigla Waite contains several supraspecific subgroups that include nominal taxa Pterygotrigla and Otohime Jordan and Starks. Studies of the genus have been complicated by a paucity of material making it difficult to obtain an understanding of the species composition of the subgenera and their relationships. All of the species share very similar meristics and body shapes, with head and cleithral spinations and colouration being the main distinguishing characters. Richards (2000) gives current information on most Pterygotrigla species, however, none of the subgenera has been appraised recently and this study of Otohime constitutes part of a wider revision of the genus under study by us. The subgenus Otohime comprises six nominal species of which most are poorly represented in collections: P. hemisticta (Temminck & Schlegel); P. spirai Golani & Baranes and P. arabica (Boulenger) (both formerly synonymized in hemisticta); P. multipunctata Yatou & Yamakawa; P. tagala (Herre & Kauffman); and P. spinosa Asano & Okamura which Eschmeyer (1998) considered to be a synonym of tagala. An additional six undescribed species were discovered in this study -one each from the western Indian Ocean, Maldives, Philippines, Coral Sea, and two species from northwestern Australia. In this paper, we provide diagnoses of all valid nominal species of the subgenus Otohime and describe six new species. The status of Prionotus alepis Alcock is also discussed, and a range extension for Pterygotrigla macrorhynchus is noted. The genus Pterygotrigla has several other subgenera that are also under study by us. The subgenus Pterygotrigla comprises an unresolved species complex, P. picta (Günther), P. andertoni Waite, P. pauli Hardy, and P. polyommata (Richardson) from southern temperate areas off Chile and around Australia and New Zealand. It possibly also includes a few species from the tropical western Pacific, P. acanthoplomate (Fowler) and a yet to be identified species. These taxa are characterized by small opercular spines, large cleithral spines, short and strong rostral spines, no nasal or antrorse spines, or spines on the bases of second dorsal-fin rays (Richards, Last and Yeung, in prep.). The remaining Pterygotrigla species are very poorly represented in collections and relationships are unresolved. Most of these, which have prominent nasal spines, very long rostral spines, and spines on the bases of second dorsal-fin rays, are widely distributed in the Indo-Pacific. It is difficult to assess if the wide variability in rostral spine morphology is due to ontogenetic changes or species differences, because so few specimens are known. These species include P. macrorhynchus Kamohara, P. hoplites (Fowler), and P. megalops (Fowler) with very long rostral spines; P. robertsi del Cerro & Lloris, P. ryukyuensis Matsubara & Hiyama, P. guezei Fourmanoir, and some undescribed species that have stout rostral spines; and P. multiocellata (Matsubara) that has an unique antrorse spine at the base of the rostral spine. P. leptacanthus (Günther) lacks dorsal bucklers and its generic placement is uncertain. The monotypic Uradia macrolepidota Kamohara with large trunk scales is considered to be the sister genus of Pterygotrigla. MATERIALS AND METHODS Counts and measurements follow Richards and Saksena (1977) and revisions made by Richards (1992). Both of those papers dealt with the genus Lepidotrigla and some additional emendations are necessary for the genus Pterygotrigla. Care must be taken in counting pectoral-fin rays because the upper two rays are often conjoined and do not appear to be separate, and the lowermost ray may be small and easily overlooked. The bases (proximal ends) of the first dorsal-fin spines are positioned between the bucklers (expansions of the pterygiophores) along the base of the fin, but no spine has been observed between the last two bucklers of this subgenus. The dorsal-fin spines are progressively smaller posteriorly, and the last one (often very short and fixed) is also included in the count. A spine has not been observed between the last two bucklers in this subgenus. The first, spine-like element of the anal fin and the first element of the second dorsal fin (which may appear spinelike) are recorded as soft rays. Caudal fin rays are difficult to count except on radiographs or cleared and stained specimens. Principal caudal-fin ray counts include those articulating directly on the hypural or parhypural bones rather than the all branched and the single, enlarged unbranched rays above and below. The principal rays are always much denser on radiographs than the secondary rays. The gill-raker counts apply to developed rakers rather than the short rudimentary rakers that are difficult to count. There are 1 or 2 small rudimentary rakers in those species without epibranchial rakers. Rudimentary rakers may also appear at the anterior end of the ceratobranchial and are always present on the hypobranchial bone. The pterygiophore interdigitation formula follows Ahlstrom et al. (1976). In counting vertebrae and determining pterygiophore interdigitation using radiographs, the first two trunk vertebrae are difficult to discern because of the heavy ossification of the cleithrum. Fortunately, the interneural space between the third and fourth neural spines always has two pterygiophores inserted there, thus serving as an excellent reference point for determining vertebrae numbers and interdigitation. The pterygiophore interdigitation is given with a virgule (/) representing neural or haemal spines and a number indicating the number of pterygiophores in the intervening interneural space. It is also difficult to differentiate trunk and caudal vertebrae, as this distinction is not clear. In radiographs that we examined pleural ribs sometimes are visible on the 12th vertebrae whereas the first two anal pterygiophores insert between the 11thand 12thvertebrae thus yielding different reference points. Consequently we report only total number of vertebrae. The condition of the cleithral spine varies from well developed and projecting from the widened area of the cleithrum, to small (often reduced to a triangular point), or absent altogether with just a widening of the cleithrum in the area where a spine would normally occur. The ‘cleithral spine width’ for all species is measured across this widened area from the anterior edge of the cleithrum to the posterior edge, whether or not a spine is present small values indicate that a spine was not present. The presence or absence of scales cannot be easily determined in some specimens due to poor preservation, but if a scale or clear evidence of a scale pocket is found in an area then it is termed scaled. We examined scale condition superficially (cleared and stained with alizarin red from a skin sample taken from the right flank below the end of the first dorsal fin) as both lateral line and body scales are small (Table 5 and Fig. 1), but with further study they may prove to have some significance. The pectoral-fin colouration and pattern is better seen in fresh specimens, as the pectoral fins of old preserved specimens are difficult to open to observe the pattern. Similarly, colouration and pattern are difficult to assess in torn vertical fins. Each membrane between the dorsal-fin spines must be carefully examined to detect presence or absence of pigment and the extent of its coverage. Measurements are taken in the horizontal (and vertical) axis (es). For example, head length is the distance between two vertical lines that are drawn at the symphysis of the premaxillary bones and the posterior edge of the opercular flap (rather than to the tip of the opercular spine as that spine length is highly variable in the genus). Snout length is the horizontal distance from the symphysis of the premaxillaries to the anterior edge of the orbit. Length of the first infraorbital (rostral) spine is the horizontal distance from the anterior tip of the longest spine to the premaxillary symphysis. The width of the first infraorbital spine is measured across the spine’s base at a point level with the premaxillary symphysis. Standard length also originates at the premaxillary symphysis and extends to the base of the caudal rays. Thus, several measurements (head length, snout length, rostral spine length, and standard length) all share a common point the premaxillary symphysis. This point is also on the same vertical plane as the point where the left and right first infraorbital bones and nasal bones converge to form a bridge between the rostral spine bases and this point is located directly above the premaxillary symphysis. Institutional acronyms are listed in the Acknowledgements section at the end of this paper and follow Leviton et al. (1985). Otohime Jordan & Starks 1907:131. Type species Trigla hemisticta Temminck & Schlegel 1843: 36 by original designation. DIAGNOSIS: Opercular spine long and slender, extending posterior to cleithrum, nasal spine absent, antrorse rostral spine absent, spines absent at the base of second dorsal-fin softrays, cleithral spine present, reduced or absent. Principal caudal rays usually 13. Number of vertebrae 25 or 27, rarely 26 or 28. Pterygiophore interdigitation: dorsal //1/2/1/1/1/1/ 1/1/1/1/1/2/1/1/1/1/1/1/1///// and ventral 2/ 1/1/1/1/1/1/1/1/1///// . SPECIES: P. hemisticta (Temminck & Schlegel) from the western Pacific from Japan to northwestern Australia, P. tagala (Herre & Kauffman) from the Philippines, P. multipunctata Yatou & Yamakawa from Japan, P. arabica (Boulenger) from the Arabian Sea, P. spirai Golani and Baranes from the northern Red Sea, P. amaokai new species from the southwestern Indian Ocean, P. hafizi new species from the Maldives, P. urashimai new species from the Philippines, P. soela new species from the Coral Sea, and P. draiggoch new species and P. elicryste new species from the continental shelf off northwestern Australian shelf. Key to the species of Pterygotrigla (Otohime) 1a. Pectoral fin with 12 connected rays ........................ 2. 1b. Pectoral fin with 13 or rarely 14 connected rays ... 4. 2a. Inner surface of pectoral-fin usually with white spots or dots near proximal part; snout relatively short, length 10-15% SL ....................................................... 3. 2b. Inner surface of pectoral fin with black pigment on membranes and rays, but lacking white spots or dots; snout relatively elongate, length 15-19% SL ..................................................................................... P. arabica 3a. Black pigment confined to membranes of inner surface of pectoral fin, several white spots of different sizes and shapes on the proximal part of fin; snout relatively short, length 10-13 % SL, trunk scales elongate, vertebrae 27 ..................................................... P. spirai 3b. Black pigment all over inner surface of pectoral fin, intense black blotch near base bordered by a diagonal row of white spots; snout moderate, length 13-15 % SL, trunk scales with scalloped edge, vertebrae 25 ......................................................................... .P. hemisticta 4a. Interspinous membranes of first dorsal fin lacking black pigment ............................................................. 5. 4b. Interspinous membranes of first dorsal fin with black pigment in the form of a blotch (rarely absent in P. elicryste and confined to small spots in P. multipunctata) ..................................................................................... 7. 5a. Inner surface of pectoral fin with one or more white, ocelli-like spots around an intense black blotch; cleithral spine moderate to small or appearing as an anglular projection; scales present on breast ............ .......................................................................... P. tagala 5b. Inner surface of pectoral fin lacking white ocelli-like spots; cleithral spine rudimentary or absent; scales maybe absent on breast ............................................. 6. 6a. Second dorsal-fin rays 12; gill-rakers on ceratobranchial 10-11; breast usually with scales ........................................................................... P. amaokai 6b. Second dorsal-fin rays 13; gill-rakers on ceratobranchial 8; breast lacking scales ..................... .................................................................... P. urashimai 7a. Black pigment on first dorsal fin partly located on first 1 or 2 interspinous membranes ........................ 8. 7b. No black pigment on first 1 or 2 interspinous membranes of first dorsal fin .................................. 10. 8a. Prepectoral area lacking scales ................................. 9 8b. Prepectoral area with scales ...................... P. elicryste 9a. Black pigment on first dorsal-fin interspinous membranes as large distinct spots on membranes 1-3, breast usually scaled, snout length 10-15% SL ......... ............................................................................ P. soela 9b. Black pigment on first dorsal-fin interspinous membranes as small indistinct spots on membranes 2-6, breast unscaled, snout length 15-16% SL ........... ............................................................. P. multipunctata 10a. Inner surface of pectoral fin usually with white ocelli-like spots; dorsal-fin spines 6-8; 9-10 bucklers .............................................................................. P. draiggoch 10b. Inner surface of pectoral fin without white ocelli-like spots; dorsal-fin spines 9; 5 bucklers .... P. hafizi (Refer also to comparison of characters in Tables 1, 2, 3, 4, 5, 6, 7) Petrygotrigla (Otohime) amaokai sp. nov. (Fig. 1E; PL.1, Fig.2; Tables 1, 2, 3, 4, 5, 6, 7) HOLOTYPE: HUMZ 73367, 121.7 mm SL, Indian Ocean, Saya de Malha Bank, 11o28’S, 061o13’E, 176 m. PARATYPES: USNM 356932, 1 (132.3 mm SL) Indian Ocean, off Kenya, 02o24’S, 41o08’E, PROF. MESIATZEV, Tr. 182, 18 June 1976, 160-165 m. USNM 356393, 2 (110.9-138.2 mm SL), Indian Ocean, Saya de Malha Bank, 11o27’S, 061o00’E, PROF. MESIATZEV Tr. 453, 10 April 1977, 156-158 m. HUMZ 72331, 1 (96.2 mm SL), Indian Ocean, Saya de Malha Bank, 11o04’S, 062o10’E, 187 m. The following paratypes were collected with the holotype: HUMZ 73363, 1 (127 mm SL), HUMZ 73364, 1 (125.7 mm SL), HUMZ 73365, 1 (130.5 mm SL), HUMZ 73366, 1 (137.6 mm SL), HUMZ 73368, 1 (138.8 mm SL). DIAGNOSIS: A species of Pterygotrigla (Otohime) without a definite cleithral spine (4-6 % SL), rudiment of the spine medial to the opercular spine; no black blotch in first dorsal fin, but red spot present in fresh material (Fig. 1); trunk scales round; prepectoral and interpelvic area naked; breast usually scaled, rarely naked; belly scaled; pectoral fin with 13 (rarely 14) connected rays, black blotch near base and membranes dark, pectoral length 35-44 % SL, longest free ray 32-36 % SL; epibranchial gill-rakers 0-1, ceratobranchial gill-rakers 10-13, short (2-3 % SL). Vertebrae 27, rarely 26. DESCRIPTION: Counts and measurements of holotype and paratypes are given in Tables 1, 2, 3, 4, 5, 6, 7. Head moderately large with short rostral spines; large nuchal spines; elongated opercular spine; sharp spine on preopercle with smaller spine below followed by an indentation and large blunt projection; no other spines on head, but head slightly rugose not smooth. Eye large and laterally positioned, slightly longer than deep; interorbital broad usually narrower than orbital width and wider than orbital depth; mouth large with fine teeth on premaxillaries, dentaries, and head of vomer. Trunk deep anteriorly, tapering posteriad with a narrow caudal peduncle. Ventral aspect not flattened as in other triglid genera. Fins moderate with caudal fin slightly lunate. Pectoral fin moderate, reaching past anal-fin origin; pelvic fin also reaching beyond anal-fin origin. First dorsal fin spine finely serrate. Trunk scales small, lacking ctenii; lateral line scales simple, tubular, with small dorsal pores (difficult to count). Scales absent from nape, prepectoral area, and interpelvic area, but present on the belly. Breast squamation variable, with only a few scales in one to several small patches (rarely completely lacking scales). Cleithral spine lacking, cleithral plate present. Four of the 6 specimens X-rayed had the last two anal pterygiophores inserted in the same interhaemal space. COLOUR IN LIFE: From colour photos of fresh holotype (HUMZ 73367) and a paratype (HUMZ 72331). Body pale pink with a deep red blotch below first dorsal on flank above pectoral fin. First dorsal fin pink without black pigment spot; few small, pale brown spots on dorsum below second dorsal fin. Iris yellow encircled with brown. Inner surface of pectoral fin with translucent distal margin; interradial membrane dark gray with black basal patch; rays lacking pigment, except for medial 4 or 5 rays covered by black blotch near base; no white, ocelli-like spots. DISTRIBUTION: Outer continental shelf off Kenya, East Africa, and on the Saya de Malha Bank some 30o east of Africa in the western Indian Ocean in depths of 156-187 m. ETYMOLOGY: Named after the eminent and recently retired Japanese scientist, Kunio Amaoka, (formerly at Hokkaido University), for his many contributions to ichthyology. REMARKS: This species is known from two localities in the western Indian Ocean. Richards (1992) reported on species of the triglid genus Lepidotrigla from this region. A continental African species, L. multispinosa Smith, also occurs across the Mozambique Channel along the coast of Madagascar, but a congener, L. alcocki Regan, is endemic to the Saya de Malha Bank. The presence of P. amaokai in Kenyan waters and at the Saya de Malha Bank and not in intervening areas is likely to reflect the paucity of deep demersal collecting in the Seychelles and Mascarene Plateau regions. Pterygotrigla (Otohime) arabica (Boulenger, 1888) (Fig.1 B – C; Pl.1, Fig. 3; Tables 1, 2, 3, 4, 5, 6, 7) Trigla arabica Boulenger, 1888: 663 (Muscat, Oman). Boulenger 1889: 245-246 + illus. Day, 1888 (description, synonymized in T. hemisticta). Steindachner, 1902: 165. Eschmeyer, 1998: 122 (list, type, validation). Trigla hemisticta (non Temminck & Schlegel): Day, 1888: 791; Day, 1889: 241; Alcock, 1890a: 197-222; Alcock 1890b: 295-311; Alcock 1896: 319; Alcock 1899:211. Prionotus alepis Alcock 1896: 313 (tentatively synonymized). Menon & Yazdani, 1968: 156; Richards & Saksena 1974: 58. Pterygotrigla picta (non Günther): Samuel 1963: 97-121. Pterygotrigla hemisticta (non Temminck & Schlegel): Richards & Saksena 1974: 57-59 (illustration, synonymy, description, discuss synonyms, distribution). HOLOTYPE: Trigla arabica, BMNH 1887.11.11.233, 1 (197.5 mm SL), Muscat, Oman, A. S. G. Jayakar. MATERIAL EXAMINED: IOES 302, 16 (86.5-115.2 mm SL) Sta. 189, off Cochin, South India, otter trawl, 138-210 m. IOES 303, 1 (97.9 mm SL) same data as IOES 302. USNM 356390, 16 (40.7-121.8 mm SL), R/V ANTON BRUUN Cr. 4B, Sta. 245A, 24o55’N, 061o10’E, 28 Nov. 1963. USNM 44426, 3 (96.0-129.2 mm SL) INVESTIGATOR, Bay of Bengal. BMNH 1910.1.31.24, 1 (61.3 mm SL) Persian Gulf, Oman, 10o W of Dubai, Coll. F. W.Townsend. 26 m. DIAGNOSIS: A species of Pterygotrigla (Otohime) with cleithral spine 7.7-11.0 % SL and ventraL to opercular spine; black blotch on interspinous membrane between first dorsal-fin spines 3-5; trunk scales elongate; prepectoral area scaled or naked; breast usually naked, rarely scaled; interpelvic area naked; belly scaled; pectoral-fin with 12 connected rays; most membranes and rays dark, with black area near base, no white spots; pectoral length 29-40 % SL; free pectoral ray 28-39 % SL; snout 15-19 % SL, orbit 11-14 % SL, epibranchial gill-rakers 1-2; ceratobranchial gill-rakers 10-15, long and exceptionally slender, 4-6 % SL. Vertebrae 27, rarely 28. DISTRIBUTION: Known from the continental shelf off Oman in the Persian Gulf and in the eastern Arabian Sea, off India, in 26-210 m. REMARKS: This species has been mistakenly synonymized with P. hemisticta, but differs from it by lacking white spots on the inner surface of the pectoral fin and having slightly more numerous (ceratobranchial modally 12-13 vs 10-11) and longer gill-rakers (2.9-6.7 % SL vs 2.6-4.3 % SL). Golani and Baranes (1997) considered that P. arabica was valid and distinct from P. hemisticta, based on Boulenger’s (1889) accounts of the two forms. Boulenger (1889) listed three characters for separating the two species (he erroneously refers to hemisticta as polysticta): development of the bony plates (bucklers) on the dorsal-fin base (actually expanded pterygiophores), orbit size, and distance between the first and second dorsal fins. However, we have found significant intraspecific variation in these characters, thus colour pattern and other features as used in the diagnoses and key are more reliable for distinguishing P. arabica and P. hemisticta. The type specimen of P. arabica is much larger than other specimens of either this species or P. spirai. This adds to the difficulty in making species comparisons based on morphometry because measurements made of the large type specimen fall well outside the ranges of all other material. Golani and Baranes (1997) mentioned that Richards and Saksena (1974) and Richards (1984) ignored some characters when comparing the type of P. arabica with P. hemisticta. However, the characters supposedly ignored do not display apparent differences when comparing specimens of similar sizes. Pterygotrigla (Otohime) draiggoch sp. nov. (Fig.1D; Pl. 1, Figs. 4-5; Tables 1, 2, 3, 4, 5, 6, 7) Pterygotrigla hemisticta (non Temminch & Schlegel): Gloerfelt-Tarp & Kailola, 1984: 118-119 (colour photo, notes). Allen & Swainston, 1990: 52-53 (range, diagnostic characters, colour illustration). Paxton et al., 1989: 456 (list, distribution). HOLOTYPE: CSIRO CA1535, 221 mm SL, NE of Monte Bello Island, Dampier Archipelago, Western Australia; 19o30’S, 116o01E to 19o31’S, 115o59’E, FRV SOELA So01/ 79/17, 3 December 1979, 142 m. PARATYPES: NTM S12931-005, 3 (56-79 mm SL), Arafura Sea, Northern Territory, Australia, R. Williams RW 9033, 16 November 1990, 124 m. Several lots from vicinity of the holotype: AMS I.22805-002, 3 (83-93 mm SL), 18o28’S, 118o15E, 1982, 156 m. CSIRO CA4196, 1 (200 mm SL), off Port Headland, 18o38.6’S, 118o02.7’E to 18o 09.0’S, 118o04.0’E, FRV SOELA So03/83/26, 6 June 1983, 182-184 m. WAM P26193, 4 (73-91 mm SL) 150 km NNW of Rosemary Island, Dampier Archipelago, 19o17’S, 116o16’E, B. Hutchins on ‘COURAGEOUS’ 16 May 1978, 170-172 m. WAM P26182, 1 (92 mm SL), Muirow Island, 21o25’S, 114o18’E, B. Hutchins on ‘COURAGEOUS’, 6 May 1978, 175-185 m. WAM P26187, 1 (208 mm SL), 55 km NW Monte Bello Island, 19o57’S, 115o13’E, B. Hutchins on ‘COURAGEOUS, 10 May 1978, 80-150 m. DIAGNOSIS: A species of Pterygotrigla (Otohime) with cleithral spine present, 5-9 % SL, ventral to opercular spine; black blotch on interspinous membranes between dorsal-fin spines 3-5, (rarely extending on 6thspine); trunk scales elongate; prepectoral area naked; breast usually scaled, rarely naked; interpelvic area naked; belly scaled; pectoral fin dark with 13 connected rays, usually with several white or light spots on lateral edge of black area and 2-3 dark spots on upper edge of fin, pectoral length 39-50 % SL; longest pectoral free ray 31-45 % SL; epibranchial gill-rakers 0-2, ceratobranchial gill-rakers 5-10, short, about 2 % SL. Vertebrae 27. DESCRIPTION: Counts and measurements of the holotype and paratypes are given in Tables 1, 2, 3, 4, 5, 6, 7. Head with short, straight rostral spines; spines slightly widened and triangular and not spread at tips; large nuchal spines extending almost to level of first dorsal spine base; opercular spine extending slightly posterior to cleithral spine; two preopercular spines, upper longer than lower; no other spines on head, although preorbital bones almost spine-like as they protrude forward anteriorly of eyes. Eye large and laterally positioned, orbit length variable, but about equal to interorbital width. Mouth large with fine teeth on premaxillaries, dentaries, and head of vomer. Trunk deep, anteriorly tapering posteriorly with narrow caudal peduncle. Ventral aspect not flattened as in other triglid genera. Caudal fin slightly lunate; pectoral fin moderate, reaching middle of anal fin; 1st free pectoral ray reaching base of anal fin; pelvic fin not reaching anal opening. First dorsal-fin spine finely serrate. Lateral-line scales of holotype with complex ossified structure (Fig. 1). Body scales with odd elongate shape (Fig. 1) and closely overlapping; scales absent from nape, prepectoral area, and interpelvic area, present on breast and belly. First dorsal-fin spines and preorbital bones showing evidence of hyperostosis on large specimens. COLOURATION: Small dark spots on dorsum below dorsal fins with a few dark spots on nape, top of head, and snout. First dorsal fin with black spot on interspinous membranes between 3rd and 5th spines. Second dorsal fin usually uniformly pale, small dark spots rarely present; caudal and pelvic fins pale. Pectoral fin with 23 dark spots on pale anterior rays; membranes between th and 6th rays dusky; membranes between 7th and 9th rays black, with few white streaks or spots on each ray; lower ray membranes pale; black area extending from near base of pectoral fin to over two thirds the fin’s inner surface; remainder of fin membranes dusky to fin margin with no discernible border along fin margin. Fresh colours: (Fig. 4 and 5) upper body and head reddish, with small dark spots. Anterior edge of pectoral fin, pink. DISTRIBUTION: Poorly known species represented by a few specimens collected from the outer continental shelf off northwestern Australia in 80-185 m. ETYMOLOGY: Due to its spiny appearance and reddish color it is named after the Welsh red dragon by combining the Welsh words draig (dragon) and goch (red). The name is treated as a noun in apposition. Pterygotrigla (Otohime) elicryste sp. nov. (Fig. 1G; Pl. 1, Figs. 6-7; Tables 1, 2, 3, 4, 5, 6, 7) Pterygotrigla hemisticta (non Temminck & Schlegel): Sainsbury et al., 1985: 104-105 (description, colour photo). HOLOTYPE: CSIRO H1022-4 (83.1 mm SL), Western Australia, off Port Hedland, 18o21’S, 118o53’E, 138 m. PARATYPES: CSIRO CA1593, 1 (65.4 mm SL), Western Australia, NE of Monte Bello Island, 19o41S, 116o12’E, FRV SOELA So04/80/18; 124 m; CSIRO CA1594, 1 (74.0 mm SL), Northern Territory, N of Bathurst Island, 10o14’S, 130o03’E, FRV SOELA So05/80/57; 124 m; CSIRO CA1595 (58.4 mm SL), taken with CSIRO CA 1594. DIAGNOSIS: A species of Pterygotrigla (Otohime) with cleithral spine present or absent (7-9 % SL when present) and ventral to opercular spine when present; interspinous membranes of first dorsal-fin sometimes with black pigment distally between spines 1-3 and spines 5-7; trunk scales crenulate, prepectoral area, breast and belly scaled; interpelvic area usually naked; outer surface of pectoral fin gray, with 13 connected rays, black area on inner side of ventral 3 rays and usually with several pale streaks on its lateral edge; pectoral-fin length 42-51 % SL, longest free ray 44-48 % SL; epibranchial gill-rakers 0-1, ceratobranchial gill-rakers 8-9, length about 3 % SL. Vertebrae 27, rarely 26. DESCRIPTION: Counts and measurements of the holotype and paratypes are given in Tables 1, 2, 3, 4, 5, 6, 7. Head large with moderate rostral spines diverging at tips; nuchal spines long extending beyond base of third dorsal spine; opercular spines extend beyond base of 4th dorsal spine; two preopercular spines, upper much longer than lower; no other spines on head. Cleithral spine shape variable: short, pungent in holotype; present on both sides in paratype CSIRO CA1593; absent on right side and plate present on left in CSIRO CA1594; and very small, hardly extending beyond plate in CSIRO CA1595. Eye very large, slightly wider than deep, laterally positioned; orbit width longer than interorbital width. Mouth large with fine teeth on premaxillaries, dentaries, and head of vomer. Trunk deep anteriorly, tapering posteriorly with narrow caudal peduncle. Ventral aspect not flattened as in other triglid genera. Fins moderate, caudal-fin lunate; pectoral-fin elongate, reaching beyond base of 3rd anal fin ray (to 9th ray in holotype), 1st free pectoral-fin ray reaching 2nd anal-fin ray; pelvic fin usually not reaching anal opening, (beyond opening in holotype). Lateral-line scales simple, tubular, without spination, with a single pore (Fig. 1). Body scales closely overlapping and without ctenii; and nearly round with 3 or 4 scalloped edges (Fig. 1); scales absent from nape, present on prepectoral area in holotype and 2 smallest paratypes, present on breast, usually absent from interpelvic area (but present on holotype), and present on belly. COLOURATION: In preservative, dark spots absent on holotype (present dorsolaterally on trunk and a few spots on top of head near base of nuchal spine of paratypes). Lower flank pale yellowish to tan. First dorsal-fin sometimes with black spots on membrane between spines 1-3 and spines 5-7; small dark spots submarginally on second dorsal-fin; pelvic and caudal fins pale. Pectoral fin with gray pigment on membranes; rays pale, except for an intense black area on ventral 3 rays, bordered dorsally with 3 to 5 white, irregularly shaped streaks. Fresh paratype (Fig. 7) with dorsal surface of snout, head and trunk reddish brown; dark spots on dorsum along fin bases and on lateral line. Possible reddish spot on first dorsal-fin margin, but membranes torn. Dark spots on second dorsal-fin. Pectoral fin with light rays and dark membranes with some reddish and dark spots on upper rays, distinctly white, irregularly shaped spots bordering black area; pale distal margin. DISTRIBUTION: Poorly known species represented by a few specimens collected from the outer continental shelf northwestern Australia in 124-138 m. ETYMOLOGY: Arbitrary combination of the first names of the mothers of the senior author’s grandchildren (Elizabeth, Crystal, and Stephania), treated as a noun in apposition. REMARKS: Known from a similar area and depth as P. draiggoch, but was not collected at the same trawl stations. Pterygotrigla (Otohime) hafizi sp. nov. (Fig.1 F; Pl.1, Fig. 8; Tables 1, 2, 3, 4, 5, 6, 7) HOLOTYPE: BMNH 1997.1.6.1, (88.7 mm SL), NANSEN Survey, 06o18’N, 073o14’E, Maldives, 25 Aug. 1983, 230 m. DIAGNOSIS: A species of Pterygotrigla (Otohime) with cleithral spine present (width about 10 % SL) and ventrad of the opercular spine; black blotch on first dorsal-fin membranes between spines 3-6; first dorsal fin bucklers 5; prepectoral area naked; breast scaled; interpelvic area naked; nape naked; belly scaled; pectoral fin with 13 connected rays, dusky with distinctive black area over rays and membranes 4-8, membranes of rays 9-11 with black pigment, remainder of fin pale including distal margin; pectoral-fin length 39 % SL, free pectoral ray 39 % SL; epibranchial gill-rakers 1, ceratobranchial gill-rakers 9, short 3 % SL; head and body with few pale spots. Vertebrae 27. DESCRIPTION: Counts and measurements of the holotype are given in Tables 1, 2, 3, 4, 5, 6, 7. Head large; rostral spines short, slightly broader at base; nuchal spines large, reaching beyond base of first dorsal-fin spine; opercular spine elongate, extending posterior of well-developed cleithral spine; preopercle with two spines, upper spine larger; no other spines on head. Eye large, laterally positioned; orbit length exceeding both orbit depth and interorbital width. Mouth very large, teeth present on premaxillaries, dentaries, and head of vomer. Trunk deep anteriorly, tapering posteriorly with narrow caudal peduncle. Ventral aspect not flattened as in other triglid genera. Fins moderate with caudal slightly lunate; pectoral fin reaching to middle of anal- fin base, first free pectoral-fin ray reaching almost to tip of pectoral fin; pelvic fin reaching to base of 2nd anal ray. First dorsal fin with leading edge of second spine finely serrate (first spine missing). Bucklers at base of dorsal fin 5. First anal-fin ray spine-like. Trunk scales small and round without ctenii, scales absent on nape, prepectoral area, and interpelvic area; present on breast and belly. Lateral line scales tubular, small spines present at dorsal opening of tube. Vertebrae 27. COLOURATION: Brown spots on head, one on anterior lower margin of orbit, few below orbit, and few on top of head. Few spots visible on trunk along lateral line, but specimen poorly preserved. Distinct black spot in first dorsal fin starting on membrane between 3rd and 4th spines, continuing posteriorly and ending midway between 6th and 7th spines. Pectoral fin lacks white ocellus-like spots; margin translucent, interradial membranes black between 2nd and 5th rays followed by large intense black area covering membranes and rays of th through 10th rays (see Fig.11). All other fins pale, lacking dark pigment. DISTRIBUTION: Known only from the holotype taken from the Maldives in 230 m. ETYMOLOGY: Named in honor of the Acting Director of the Marine Fisheries Section of the Maldives, Mr. Ahmed Hafiz. REMARKS: This species, though represented by only one specimen, is unique within the subgenus in lacking dark spots on the trunk and only having 5 bucklers at the base of the first dorsal fin, possessing scales on the breast, and a distinctive pectoral fin pigmentation. It was collected in deeper water than most other members of the subgenus in a region that has not been well surveyed. Only two trigloids (both peristediids) Satyrichthys investigatoris (Alcock) and an unidentified Satyrichthys species, have been listed from the Maldives (Randall and Anderson, 1993; Adam et al. 1998). Anderson et al. (1998) in their recent new records from the Maldives did not list the P. hafizi type. Pterygotrigla macrorhynchus (Kamohara) should also be added to regional fauna lists based on a specimen provided by N. Merrett from the Maldivian collections in the BMNH. Pterygotrigla (Otohime) hemisticta (Temminck and Schlegel, 1843) (Fig.1A; Pl. 1, Fig. 9; Tables 1, 2, 3, 4, 5, 6, 7) Trigla hemisticta Temminck & Schlegel, 1843, Pisces, in Siebold, Fauna Japonica: 36 + Pl. 14. Boeseman, 1947: 46 (description, type designation). Eschmeyer, 1998: 718 (list, types, validation). Prionotus alepis Alcock, 1896: 313 (tentatively synonymized). Menon & Yazdani, 1968:156. Richards & Saksena, 1974: 58. Eschmeyer, 1998: 71 (list, types). Otohime hemisticta: Jordan & Richardson, 1908: 658-659 (synonymy, description). Pterygotrigla hemisticta: Jordan et al., 1913: 289-290. (list, illustration). Matsubara & Hiyama, 1932: 10-11(synonymy, description). Kuronuma, 1939: 251-253 (synonymy, description). Kamohara 1952: 72 (list). Kamohara, 1958: 59 (list). Kamohara. 1964: 78 (list). Masuda et al., 1975: 147,343 (colour photo, meristics, habitat, range). Ochiai and Yatou, 1984: 334, pl.300-H (description, colour photo). Shen, 1984: 32, pl.32 (colour photo, meristics, habitat). Chen and Shao, 1988: 134135, 138 (synonymy, diagnosis, illustration). Paxton et al., 1989: 456 (list, distribution). Froese et al., 1996: 237 (list). Golani & Baranes, 1997: 185 (comparisons). Richards (in press) (list, illustration, identification characters) TYPE MATERIAL: Not examined. Boeseman (1947) designated RMNH 695 (stuffed specimen 265 mm) as lectotype and listed RMNH 501 (preserved specimen 230 mm) as a paralectotype MATERIAL EXAMINED: USNM 57569, 1 (181 mm SL) Japan. KU uncataloged, 2 (133.4-156.6 mm SL), Taiwan. CSIRO H2548.11, 2 (140.0-169.0 mm SL), Indian Ocean, Western Australia, NW of North West Cape, 21o44.7’S, 113o52.3E to 21o44.5’S, 113o52.5’E, FRV SOUTHERN SURVEYOR, SS01/91/8, 24 January 1991, 320 to 290 m. CSIRO H4031-28. 1(92.7 mm SL) INFO. NTM S-2589-016, 1 (231 mm SL), Indian Ocean, Western Australia off Rowley Shoals, NW Shelf, W. Houston, sta 85-16, 2 November 1985, 430 m depth. NTM S-12609-015, 1 (245 mm SL), Indian Ocean, Western Australia off Rowley Shoals, NW Shelf, W. Houston, sta 85-31, 7 November 1985, 420 m. DIAGNOSIS: A species of Pterygotrigla (Otohime) with cleithral spine present (width 8-10 % SL) and ventrad of opercular spine; black blotch in first dorsal-fin on interspinous membranes between spines 3-6; trunk scales with scalloped edges; prepectoral area scaled or naked; breast scaled; interpelvic area naked; belly scaled; pectoral fin dark with black base and diagonal band of separate white spots, 12 connected rays; pectoral fin 32-36 % SL; free pectoral ray 29-37 % SL; epibranchial gill rakers 1; ceratobranchial gill rakers 10-12, long and slender, 3-4 % SL. Second dorsal-rays 11 and only 5 pterygiophores after the double pterygiophore supporting the second dorsal-fin. Vertebrae 25. Large specimens exhibit hyperostosis. DISTRIBUTION: Widely distributed in the western Pacific from Japan to Australia (Richards, in press) where it occurs in shallow coastal waters (20 m) as well as on the upper continental slope (220-420 m). REMARKS: This species is brightly coloured with a red dorsum covered with dark spots and a distinctive row of white spots on a black background on the inner surface of the pectoral fin (Fig. 8). Richards and Saksena (1977) mistakenly synonymized it with P. arabica as have other authors (Day, 1888; Alcock, 1890, 1899; Richards, 1984) and is likely to be closely allied to P. spirai. P. hemisticta differs from P. arabica and P. spirai in having fewer vertebrae (25 vs 27-28). It also differs from P. arabica in pectoral fin coloration (no white spots on inner surface of pectoral fin), a marginally smaller eye and cheek height, and shorter gill-rakers (Tables 1-2). P. spirai has fewer white spots on the inner surface of the pectoral fin that are not aligned diagonally, a shorter snout, and more elongate trunk scales. The reduced number of vertebrae (25) was found in all specimens that were X-rayed andthe specimens were from Japan, Taiwan, and Australia. This is the lowest number of vertebrae for any species in the Family Triglidae. Large specimens from Australia display hyperostosis with swollen head bones (infraorbitals, frontals, opercles) and spines (nuchals, opercles, and cleithrals). Hyperostosis in triglids is uncommon having been observed in one Prionotus species (Smith-Vaniz et al., 1995) and a few other Pterygotrigla (Richards, pers. obs.). Pterygotrigla (Otohime) multipunctata Yatou and Yamakawa, 1983 (Tables 1, 2, 3, 4, 5, 6, 7) Pterygotrigla multipunctata Yatou and Yamakawa 1983: 217-220. Eschmeyer 1998: 1133 (list, types). HOLOTYPE: NSMT-P 21409, (151 mm SL). Japan, Tosa Bay, Mimase, Kochi City. 4 April 1966. PARATYPE: BSKU 50036 (now at KSHS), 1 (88 mm SL) Japan, Tosa Bay, Kochi City, Mimase. 25 February 1966. DIAGNOSIS: A species of Pterygotrigla (Otohime) without a defined cleithral spine, remnant medial of the opercular spine; several small dark spots but no black blotch in first dorsal fin; prepectoral, breast, and interpelvic areas naked; belly scaled; pectoral fin with black blotch, membranes dark, 13 connected rays; pectoral length 3544 % SL, longest free ray 30-36 % SL; epibranchial gill-rakers 1, ceratobranchial gill-rakers 9-10, short (less than 3.5 % SL). Vertebrae 27. DISTRIBUTION: Known only from Tosa Bay, Japan. REMARKS: This poorly represented species is known only from the type material collected in Japan. Counts and measurements are given in Tables 1, 2, 3, 4, 5, 6, 7. Pterygotrigla (Otohime) soela sp. nov. (Figs. 10-11, Tables 1, 2, 3, 4, 5, 6, 7) Pterygotrigla tagala (non Herre & Kaufman): Del Cerro and Lloris 1997: 119-120 (brief description, listed). HOLOTYPE: CSIRO H580-02, 97.8 mm SL, Pacific Ocean, Coral Sea, off Cairns, Queensland, Australia, 17o35.3’S, 149o56.9’E to 17o33.8’S, 149o52.9’E, FRV SOELA SoO6/ 85/65, 302 m. PARATYPES: CSIRO H769-02, 1 (91.1 mm SL), Pacific Ocean, Coral Sea, E of Flinders Reef, Queensland, Australia, 17o32.8’S, 149o46.2E to 17o27.7’S, 149o46.5’E, FRV SOELA SoO6/85/64, 3 December 1985, 338-348 m. The following paratypes were collected mainly by the Research Vessel CORIOLIS from the Coral Sea, Pacific Ocean, on the western side of Chesterfield Islands, Bellona Plateau, and a nearby unnamed plateau south of the Bellona Plateau: MNHN 1995-498, 1 (95.3 mm SL), MUSORSTOM 5: Stn. CP 276, 24o48.9’S, 159o40.9’E, beam trawl, 9 October 1986, 259-269 m. MNHN 1995-499, 5 (93.6-99.9 mm SL), MUSORSTOM 5: Stn. CP 312, 22o17.2’S, 159o24.8’E, beam trawl, 12 October 1986, 315320 m; MNHN 1995-500, 2 (91.5-95.4 mm SL), MUSORSTOM 5: Stn. CP 268, 24o44.7’S, 159o39.2’E, beam trawl, 9 October 1986, 280 m; MNHN 1995-501, 1 (99.0 mm SL), MUSORSTOM 5: stn. CH 271, 24o48.2’S, 159o34.6’E, otter trawl, 9 October 1986, 250-276 m; MNHN 1995-502, 2 (92.8-104.9 mm SL), MUSORSTOM 5: Stn. CP 316, 22o25.1’S, 159o24.0’E, beam trawl, 13 October 1986, 330 m; MNHN 1995-503, 3 (91.1-110.1 mm SL), MUSORSTOM 5: Stn. CP 351, 19o33.1’S, 158o36.9’E, beam trawl, 17 October 1986, 290-310 m; MNHN 1995-504, 2 (90.4-102.4 mm SL), MUSORSTOM 5: Stn. CP 319, 22o24.4’S, 159o16.5’E, beam trawl, 13 October 1986, 320325 m; and MNHN 1995-505, 15 (74.7-108.9 mm SL), CHALCAL 1: Stn. CH 2, 22o34.4S, 159o17.4’E, otter trawl, 28 July 1984, 330 m. DIAGNOSIS: A species of Pterygotrigla (Otohime) with cleithral spine reduced to angular projection lying mediad of opercular spine, cleithrum width (3-6 % SL); black blotch on interspinous membranes between first three dorsal-fin spines; trunk scales with scalloped edges; prepectoral area naked; breast usually scaled; interpelvic area naked; belly scaled; black blotch on pectoral fin distinctive, separated from dusky membranes; 13 connected pectoral-fin rays; pectoral-fin length 39-61 % SL, free ray 27-45 % SL; epibranchial gill-rakers 1 (rarely 0 or 2), ceratobranchial gill-rakers 9-13 (rarely 8), short (2-4 % SL). Vertebrae 27. DESCRIPTION: Counts and measurements of the holotype and paratype are given in Tables 1, 2, 3, 4, 5, 6, 7. Head moderate, rostral spines slightly diverging laterally at tip, curved slightly; nuchal spines large, extending posteriad to a point below 3rd dorsal spine; opercle spine elongate, extending to a point below 4th dorsal spine, much longer than orbit diameter; two preopercle spines, upper longer than lower; no other spines on head except small spine at end of bony ridge behind eye in small specimens. Cleithral spine reduced to angular projection, lying mediad of opercular spine; exposed portion of cleithrum barely visible beyond opercle, sometimes visible as small, blunt spine. Eye nearly round and very large, laterally positioned; orbit length greater than interorbital width. Mouth very large with fine teeth on premaxillaries, dentaries, and vomer. Trunk deep anteriorly, tapering posteriorly, caudal peduncle narrow. Ventral aspect not flattened as in other triglid genera. Caudal fin lunate; pectoral fin reaching midway on anal-fin base, free pectoral-fin ray reaching mid-anal fin; pelvic fin reaching anus. Scales on body deeper than long with scalloped indentations but lacking ctenii; lateral-line scales simple, tubular, without spines. Scales absent from nape, prepectoral area, and interpelvic area, usually present on breast (these scales may be small, and difficult to discern), pronounced on belly. COLOURATION: In preservative, body and head pale, usually lacking dark spots, (if present they are usually discernible along and above lateral line). Snout, upper part of head, lower jaw, anterior upper jaw, first dorsal-fin base and area of trunk below first dorsal fin pale brown. Pale brown on caudal-fin base. Cheek, end of premaxillary, preopercle, interopercle, lower opercle, lower flank silvery white (silvery whitish area in some paratypes extending dorsally on flank behind brownish area, lost in others). First dorsal fin with intense black pigment on membranes between first three spines (not apparent where fin membranes torn). Pectoral fin with pale margin; intense black pigment over lower middle rays and membranes, separated from remainder of fin by a pale area without white spots, remainder of fin with brownish gray pigmented membranes and light rays (see Fig. 11). Second dorsal and anal fins uniformly pale. DISTRIBUTION: Known from the SW Pacific. Along the upper continental slope off Queensland, Australia, and near the Chesterfield Islands in the Coral Sea in 250-348 m. ETYMOLOGY: Named after the former CSIRO research vessel R/V SOELA. Expeditions from this vessel have provided the basis of our knowledge of tropical Australian deep-sea fishes. It is treated as a noun in apposition. REMARKS: A pigment spot on the anterior part of the first dorsal fin and the pectoral-fin pigmentation are unique within the subgenus. Its very large eye possibly reflects a preference for deep-water habitats or is due to the young age (small size). Pterygotrigla spirai Golani & Baranes (Tables 1, 2, 3, 4, 5, 6, 7) Pterygotrigla sp. Baranes & Golani, 1993: 305,318(Eilat, Gulf of Aqaba). Pterygotrigla spirai Golani & Baranes, 1997: 185-195 (Eilat, Gulf of Aqaba). Eschmeyer 1998: 1594 (list, types). Pterygotrigla hemisticta (non Temminck & Schlegel): Randall, 1996: 115 (description, colour photo). Khalaf et al., 1996: 403-404 (description, photo). MATERIAL EXAMINED (all from Gulf of Aqaba at Eilat; first two lots paratypes): HUJ14002, 8 (106.2-147.2 mm SL); HUJ 17578, 3 (78.8- 157.8 mm SL); HUJ17994, 1 (131 mm SL); HUJ18347, 1 (138 mm SL); HUJ 18346, 2 (141-150 mm SL); HUJ 18348, 1 (130 mm SL); BPBM 31858, 1 (116 mm SL). DIAGNOSIS: A species of Pterygotrigla (Otohime) with cleithral spine 8.3-11.6 % SL ventral to opercular spine; black blotch on interspinous membrane between dorsal- fin spines 4-6; prepectoral area, breast, and interpelvic area naked; belly scaled; pectoral fin with 12 connected rays, usually 8 pectoral-fin membranes dark, with black area near base, and usually several white dots of different sizes and shapes on the proximal part; pectoral length 27-34 % SL; free pectoral ray 31-39 % SL; snout short 10- 13 % SL; orbit large 12-15 % SL; 1 epibranchial gill-raker plus 1-2 rudiments, ceratobranchial gill-rakers 10-13. Vertebrae 27. REMARKS: This recently described species has been found only in the Gulf of Aqaba at Eilat, Israel. It is presumably the sister species of P. arabica, but is separable using the colouration and snout length characters noted in the key. Golani and Baranes (1997) remark that the ‘wide and roundish’ internuchal area is diagnostic, but we did not note this difference in the material examined. White spots were not visible on the inside of the pectoral fin on many of the specimens examined by us. The presence of white spots, though few, possibly place it closer to a P. hemisticta ancestor as it is isolated in the northern Gulf of Aqaba. Pterygotrigla (Otohime) tagala (Herre and Kauffman, 1952) (Fig. 12, Tables 1, 2, 3, 4, 5, 6, 7) Otohime tagala Herre and Kauffman 1952: 27-28 (original description). Eschmeyer 1998: 1645 (list, types, validation). Pterygotrigla spinosa Asano and Okamura, 1963: 49-52 (original description). Eschmeyer 1998: 1591 (list, types, synonym of P. tagala). Pterygotrigla tagala. Richards (in press) (list, illustration, identification characters). Pterygotrigla tagala (non Herre & Kauffman): Del Cerro and Lloris 1997: 119-120 (brief description, listed). MATERIAL EXAMINED: Holotype: P. tagala: USNM 202506, (96.7 mm SL) Philippine Islands, Luzon, outer Manila Bay, 117 m. Paratypes: P. tagala : USNM 202566, 1 (96.3 mm SL), Philippine Islands, Luzon, south entrance to Manila Bay, 6 mi. SE of Monja Island, off Cavite Province September 1947, 99 m [originally UW 11516]. USNM 202565, 3 (79.3-93.1 mm SL) collected with the holotype. UW 11514, 5 (78.7-97.8 mm SL) South China Sea, near Fortune Island off coast of Batangas Province, Philippine Islands, 2 October 1947, 119 m. Two specimens tentatively identified as paratypes, USNM 202567, in the original description are described as new below. Holotype: P. spinosa: FAKU S526, (95.2 mm SL) China, Tonking Bay, 17o12’N, 108o40E, 95m. Paratypes: P. spinosa: FAKU S525, 1 (73.3 mm SL) and FAKU S315, 1 (65.0 mm SL) taken with the holotype. DIAGNOSIS: A species of Pterygotrigla (Otohime) with cleithral spine reduced to short, subtriangular spine (width 5-9 % SL) and ventrad of the opercular spine; no black blotch in first dorsal fin; trunk scales with scalloped edge; prepectoral, breast, belly, and interpelvic areas scaled; pectoral fin inner surface with black blotch, bordered with pale to whitish spots, remainder of fin dusky rather than black, rays and margin pale, 13 connected rays; pectoral fin length 39-51 % SL, longest free ray 39-52 % SL; epibranchial gill-rakers 0-1, ceratobranchial gill-rakers 8-12, short (3-4 % SL). Vertebrae 27, rarely 28. DISTRIBUTION: Known from the continental shelf in the region of the South China Sea. Specimens have been taken off Luzon, northern Philippines, and in Tonking Bay, southern China, in 95-119 m. REMARKS: Eschmeyer (1998) listed P. spinosa as a synonym of P. tagala and we concur. Pterygotrigla (Otohime) urashimai sp. nov. (Fig. 1H; Pl.1, Fig.13; Tables 1, 2, 3, 4, 5, 6, 7) Plate 1: Fig. 2, 3, 4, 5, 6, 7, 8 Otohime tagala (non Herre and Kauffman 1952: 28 (two tentatively listed paratypes differentiated). HOLOTYPE: USNM 202567, (93.6mmSL). Philippine Islands, Luzon, off the coast of Cavite Province, south entrance to Manila Bay, 119 m. PARATYPE: USNM 356931, 1 (82.3mmSL) taken with the holotype. DIAGNOSIS: A species of Pterygotrigla (Otohime) without a defined cleithral spine (width 5 % SL), rudiment of spine medial of opercular spine; no black blotch on first dorsal fin; trunk scales with scalloped edge, prepectoral, breast, and interpelvic areas naked; belly scaled; pectoral fin inner surface with black blotch, membranes dusky rather than black, 13 connected rays; pectoral-fin length 42-46 % SL, longest free ray 38-39 % SL; epibranchial gill-rakers 1-2, ceratobranchial gill-rakers 8, short (3 % SL). Vertebrae 27. DESCRIPTION: Counts and measurements of the holotype and paratype are given in Tables 1, 2, 3, 4, 5, 6, 7. Head large; rostral spines short, straight, not spread at tips; large nuchal spines; opercular spine elongate, slightly longer than orbit diameter; two short preopercular spines; no other spines on head. Cleithral spine absent; posterior edge of cleithrum rounded, without ridge. Eye large, laterally positioned; orbit width equal to interorbital width. Mouth very large with fine teeth on premaxillaries, dentaries and head of vomer. Trunk deep anteriorly, tapering posteriorly, caudal peduncle narrow. Ventral aspect not flattened as in other triglid genera. Fins moderate, caudal fin slightly lunate; pectoral fin moderate, reaching base of 9th anal ray, 1st free pectoral-fin ray reaching base of 7th anal ray; pelvic fin reaching base of 2nd anal ray. First dorsal-fin spine finely serrate. Scales of paratype small with posterior scalloping, slightly longer than deep, lacking ctenii. Lateral line scales tubular, opening and tube subequal in width. Scales absent from nape, prepectoral area, breast, and interpelvic area; scales present on belly. COLOURATION: Small dark spots scattered on upper half of body and head, two rows of dark spots on second dorsal fin. Inner surface of pectoral fin with intense black blotch near base, remainder of fin dusky rather than black, except for pale margin and rays, no white ocellus. DISTRIBUTION: Known from two specimens collected from the continental shelf off Manila, Luzon, in the northern Philippines in 119 m. ETYMOLOGY: Urashima is a hero of the Japanese folktale in which Otohime (the goddess of fishes) is also an important character. REMARKS: In the original description of P. tagala, Herre and Kauffman (1952) noted that two of their paratypes differed slightly from the other types, but they retained them, somewhat tentatively, in the type series. One of us, Yato, noticed several additional differences when he examined the type series at the USNM. The lateral-line scales and body scales are identical to those of P. tagala and P. soela new species, but other characters in the diagnosis and key clearly separate P. urashimai from these species. This species could be sympatric with P. tagala as only only a few meters in depth separate the type locations. Incertae sedis Pterygotrigla alepis (Alcock) Prionotus alepis Alcock 1889: 303-304, Pl. 22, Fig.9 (original description). Alcock 1896: 319 (listed, possible synonym of Trigla hemisticta Temminck and Schlegel). Eschmeyer 1998: 71 (list, types, references). Exolissus alepis. Jordan 1923: 217. (Designated type species of Exolissus by original designation, also monotypy). REMARKS: Alcock (1896) suggested that his Prionotus alepis might be the young of Pterygotrigla hemisticta. A comparison of the type description and illustration of the 36.5 mm SL or TL? holotype with similarily sized, juveniles of P. hemisticta (46.8 mm SL, 58 mm TL) revealed some differences. In the type of P. alepis the pectoral fin reaches the end of the anal-fin base (reaching midway along anal-fin base in P. hemisticta), the opercular spine is short (rather than long), the pectoral fin is truncate (rather than pointed), the dorsum lacks spots (rather than being spotted). The lateral line is incomplete with 16 tubes visible (rather than complete), and teeth are present on the vomer and palatines (rather than confined to vomer). Prionotus alepis is either the young of another Pterygotrigla species or is valid. More data are needed before its status can be clarified. Juvenile stages of most species are rare in collections and a large size series of each species is needed to resolve early life history stage identification. Jordan (1923) erected a new genus Exolissus for this species based solely on the description, which noted that it lacked scales. Apparently, Jordan failed to realize that this description was based on a small juvenile of a species far removed from areas where Prionotus are found. DISCUSSION Otohime is represented by 11 species of generally colourful triglids characterized by elongated opercle spines, lack of nasal, antrorse rostral, and second dorsal-fin ray base spines, and a tendency for reduction of the cleithral spine. The reduction of the cleithral spine in several species is diagnostic and possibly indicates relationships between them. Three of the species have 12 connected pectoral-fin rays and the remainder have 13. The colour patterns of the inner surface of the pectoral fin and the first dorsal-fin membranes are species specific and probably play a behavioural role. It is very difficult and next to impossible to discern sex from gross visual examination because the specimens are small, many are old and poorly preserved. Sexual differences may account for some intra-specific differences, but many more and better preserved specimens are needed to allow se differentiation. Morphometric data summarized in Tables 1 and 2 provide some evidence of shape differences between Otohime species. However, because of the small sample sizes and ontogenetic variability within the group, these trends cannot be unraveled using robust statistical analyses. Of the 11 species, 4 are known from 4 or fewer specimens. Nevertheless, some interspecific differences are evident. For example, where sample sizes are substantial (n>14) and the data sets include both juveniles and adults (i.e. P. arabica and P. draiggoch), morphometric differences between species were clear: premaxillary or upper jaw length 18-23 % SL in P. arabica vs. 13-17 % SL in P. draiggoch; gill raker length 2.9-6.7 % SL in P. arabica vs. 1.7-2.1 % SL in P. draiggoch. Other characters, such as pectoral fin size could be useful (length 29-40 % in P. arabica vs. 39-50 % SL in P. draiggoch). However, as the pectoral fin’s length varies greatly with growth, data for a range of growth stages is needed to provide an adequate diagnosis. The morphometric comparisons in Table 2 show similarities in some ratios and point to possible differences between others. For example, P. draiggoch and P. elicryste occur in the same region and P. tagala and P. urashimai are also sympatric. Clear differences in pigmentation and squamation separate these species but they are very similar morphologically. Apart from pectoral-fin ray counts, the species of Otohime are meristically similar (Tables 3, 4 and 7). All spines and rays are counted and in many instances the first anal-fin ray and the first second dorsal-fin ray appear to be spine-like, but in both cases they are treated as rays. P. hemisticta is a wide-ranging species and has fewer vertebrae (fewest of any triglid) and dorsal and anal-fin rays. The very low buckler count found in P. hafizi may be an individual anomaly (reminiscent of the buckler count and illustration noted by Boulenger 1889), but no abnormalities were visible in that area or in the radiograph. Additional material is needed before a final determination is possible. The connected, pectoral-fin ray counts do reveal specific differences. P. draiggoch has the fewest ceratobranchial rakers, but there is no clear separation within the subgenus. The breast was listed as scaled even if only a small patch was present. The prepectoral area was scaled in only a few species (P. tagala and P. elicryste and some specimens of P. hemisticta and P. arabica) and this is an unusual character of the genus. The shapes of the body scales and structure of the lateral-line scales appear to be useful for distinguishing. The body scales of P. hemisticta, P. soela, P. tagala, P. urashimai and P. elicryste have scalloped posterior margins; those of P. hafizi, and P. amaokai are circular; those of P. draiggoch, P. spirai, and P. arabica are elongate with a non-symmetrical protruberance. The lateral-line scales are simple tubes in P. tagala, P. urashimai, and P. soela, while P. hemisticta, P. arabica, P. spirai, P. elicryste, P. hafizi, and P. amaokai also have simple tubes with offset pores. The lateral-line scales of P. draiggoch exhibit a complex structure with small spines. Scales of P. multipunctata were unavailable for comparison. The pattern of pterygiophore interdigitation showed some variation for those pterygiophores that support second dorsal rays. This variation is given in Table 7. In addition P. hemisticta has fewer pterygiophores because of the low numbers of vertebrae and P. amaokai was unique in having a majority of X-rayed specimens with the last two anal pterygiophores inserted in the same interhaemal space. The common vertebrae number is 27, but P. hemisticta had a reduced number (25) and this was consistent throughout its wide geographic range. P. amaokai and P. elicryste each had one specimen with 26 vertebrae, remainder 27, and P. arabica and P. tagala had a few specimens with 28. Principal caudal rays for most species are 13 with a few exceptions of 14 or 15 as shown in Table 7. Colour patterns of the fins were particularly useful in separating the species (Table 6). The pattern on the inner side of the pectoral fin (upper surface when the fish swims) exhibits interspecific variation in the extent of dark pigmentation, and the presence or absence and configuration of white spots. In the species group with 12 connected pectoral rays, P. spirai was interspecifically variable in either lacking or having white spots which makes it difficult to distinguish from P. arabica (always lacking spots) using this character. In contrast, P. hemisticta always has prominent series of white spots across the fin. Again, this must be used cautiously due to small sample sizes. We did not attempt to assess the polarity of these characters so no cladistic study was made. It is premature to hypothesize on the relationships of this group until all the species of Pterygiotrigla have been studied. ACKNOWLEDGEMENTS We thank the following individuals for providing access to their museums or the loan of specimens: K. Amaoka (HUMZ), S. Jewett (USNM), H. J. Walker (SIO), Y. Shcherbachev (MMSU), P. J. P. Whitehead (BMNH), B. Hutchins (WAM), J. Paxton, J. Leis, and M. McGrouther (AMS), B. Russell and R. Williams (NTM), K.-T. Shao (THUP), N. Merrett and A.-M. Woolger (BMNH), B. Serét and P. Pruvost (MNHN), D. Golani (HUJI), C. R. Robins (KU), I. Nakamura (FAKU), and A. Graham (CSIRO). L. W. Knapp provided specimens and a photograph from the Smithsonian Oceanographic Sorting Center. B. B. Collette of the National Marine Fisheries Service provided assistance during USNM visits and X-ray service (assisted by L. Willis). W. A. Courtenay, Jr., Florida Atlantic University, provided X-ray service. A. Roberts kindly provided advice on Welsh grammar. J. Javech assisted with artwork and photographs and D. L. Jones assisted with electronic formatting. D. C. Gledhill, M. Gomon, D. Golani, and P. Heemstra reviewed the manuscript and provided many helpful comments. REFERENCES
The following images related to this document are available:Photo images[sm03002t4.jpg] [sm03002f8.jpg] [sm03002f4.jpg] [sm03002f12.jpg] [sm03002f7.jpg] [sm03002t2.jpg] [sm03002t7.jpg] [sm03002f11.jpg] [sm03002f10.jpg] [sm03002t5.jpg] [sm03002f2.jpg] [sm03002f1.jpg] [sm03002f13.jpg] [sm03002t3.jpg] [sm03002f6.jpg] [sm03002f3.jpg] [sm03002t1.jpg] [sm03002f5.jpg] [sm03002f9.jpg] [sm03002t6.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}