 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Smithiana, Publications in Aquatic Biodiversity, Bulletin 5, August, 2005, pp. 1-28 A review of the triplefin fish genus Enneapterygius (Blennioidei: Tripterygiidae) in the western Indian Ocean, with descriptions of four new species Wouter Holleman 1 1 South African Institute for Aquatic Biodiversity, Private Bag 1015, Somerset Street, Grahamstown, 6140, South Africa. E-mail: W.Holleman@ru.ac.za. Code Number: sm05002 ABSTRACT The fishes of the tripterygiid genus Enneapterygius of the western Indian Ocean (excluding Sri Lanka) are reviewed. Four new species, E. elaine, E. gruschkai, E. genamaculatus and E. kosiensis are described and several species are redescribed. Enneapterygius elaine is known only from Rodrigues, E. gruschkai, a medium-sized species, is known from the Comoro Islands, Mauritius, St Brandon Shoals and the Chagos Archipelago; E. genamaculatus is known only from St Brandon Shoals and E. kosiensis is known only from the northern KwaZulu-Natal coast. The first and last two species are small species, less than 23 mm SL. A key is provided for the 17 species recognised in the region. INTRODUCTION Fishes of the genus Enneapterygius are characteristically small, most species being less than 25 mm long. They are generally coral reef inhabitants, their colouring and small size rendering them cryptic. The advent of scuba, ichthyocides and more sophisticated underwater photographic equipment and techniques have resulted in more of these small fishes being collected and described. However, their colours fade rapidly in preservative, leaving bottles of small straw-coloured fishes with little to distinguish one species from the next. Meristically and morphometrically many species are very similar, and consequently long-preserved specimens are difficult to identify and often relegated to the back of shelves. The taxonomic history of the family - and hence the genus - has been checkered. Rosenblatt, in an unpublished thesis, made the first - and to date only revision of the family in 1959. Some of the species descriptions took decades to appear in print, often under someone else’s name. Clark (1980) made the first substantial regional revision, the tripterygiid fishes of the Red Sea; Hansen (1986) revised Helcogramma; Hardy addressed the taxonomy of the diverse New Zealand fauna (e.g. Hardy 1984, 1986, 1987), but, it was not until 1994 that the next major revisions saw the light of day: Shen & Wu’s revision of the tripterygiid fishes of Taiwan and Fricke’s Tripterygiid fishes of Australia, New Zealand and the southwestern Pacific Ocean. The most recent is Fricke’s (1997) revision of the tripterygiid fishes of the western and central Pacific Ocean. This review of western Indian Ocean (WIO) species of Enneapterygius is not comprehensive in that much available material has not been examined. Its main purpose is to provide information to identify newly collected specimens, as well as preserved material that has lost all coloration. The revision is based on material from the St Brandon Shoals (collections of Springer in 1976), the Chagos Archipelago (collections of Winterbottom and Emery in 1979), from the Comoro Islands (collections of Winterbottom et al. in 1988) and material available at the South African Institute for Aquatic Biodiversity. This revision has also benefited considerably from Fricke’s 1994 and 1997 revisions and from Randall’s more recent work in Oman and the Arabian Gulf (Randall, 1995). METHODS AND MATERIALS Measurements were taken as in Hubbs and Lagler (1958); fin element counts follow Springer (1968). Several measurements used by me in earlier papers (e.g. Holleman, 1982 & 1986) have not been included here: measurements of snout and maxilla length in such small species using a standard dial caliper are too inaccurate and variable to be of statistical significance or taxonomic value. Snout profile was also not measured, again because the method I devised is too inaccurate for fishes this small. Similarly, vertebral counts have not been made, except for the newly described species, because they are not readily available to those who would identify the species. Pectoral-fin ray counts are given as total number: (from the top) number of simple rays, number of divided rays, number of simple rays. In all tripterygiids (except the free-swimming Obliquichthys) the lowermost 5-7 pectoral-fin rays are thickened and used as support additional to the pelvic fins. The caudal fin of tripterygiids has 7 dorsal and 6 ventral segmented, principal caudal-fin rays, of which the uppermost and lowermost 2 or 3 are simple and the remainder bifurcate, and between 3 and 8 procurrent rays dorsally and ventrally. In a few specimens one (seldom two) notched scales occur anterior to the beginning of the notched segment of the lateral line, just below the end of the anterior, pored segment, and are not included in the count. Including these and the two or three un-notched scales would result in very variable counts. In most species the scales are highly deciduous and, unless specimens are collected with care, most of the scales are lost in collecting nets. Total lateral scales are counted from the last notched lateral line scale to the pectoral-fin axil. Transverse scale counts are taken from the base of the anterior half of the second dorsal fin to the pored segment of the lateral line, and below it to the base of the anal fin. Hansen (1986) included mandibular pore patterns in her Helcogramma species descriptions, finding that they were often diagnostic for species. Williams & McCormick (1990) followed suit and pore patterns are used in this revision as well. They are given as the number of pores on the one side + the number at the symphysis + the number on the other side. Two pores at the symphysis may in some individuals join to form a single, wide oval pore rather than two distinct pores. Where this is relatively common in a species the symphysial pore is indicated 1/2. Live coloration is usually diagnostic for tripterygiid species. In all species that I am aware of and of which the live colours are known, males are more brightly coloured than females, very often with much black on the head, chest and pectoral-fin bases. In preserved specimens melanophore patterns are often distinctive and diagnostic, but these too tend to fade after extended periods in alcohol, particularly where specimens are exposed to light. The western Indian Ocean as dealt with in this revision includes the Red Sea, but excludes Sri Lanka. Sufficient material for this island was not available at this time. Table 1 summarises morphometric and meristic data for the 17 species. The following institutional abbreviations are used: AMS - Australian Museum, Sydney TAXONOMIC ACCOUNTS Genus Enneapterygius Rüppell, 1835Enneapterygius Rüppell, 1835: 2; type species E. pusillus Rüppell, 1835, by monotypy. DESCRIPTION. Small fishes with fusiform bodies, less than 25 mm SL (14 of the 16 WIO species - there are Pacific species that are considerably larger). First dorsal fin with 3 spines; second with 11-14 spines; third with 8-11 simple rays, except the last which is divided to its base. Anal fin with a single, short spine and 15-22 simple rays, except for the last which is divided to its base. Height of first dorsal fin is about the same height as second or considerably taller. Pelvic fins with one short, hidden spine and two slender, simple rays which may be united by a membrane for part of the length of the shorter ray. The supratemporal sensory canal is either U-shaped with a pore at each posteromedial corner of the canal, curving around the first dorsal fin, or variously curved (Fig. 1). The mandibular canal has either one or two pores at the jaw symphysis and 3-5 pores either side (except for 2 species). Exposed edge of posttemporal bones with fine serrations. Body with ctenoid scales; nape naked or scaled; abdomen naked or with thin cycloid scales (which are difficult to see) in three species. Lateral line discontinuous, an anterior segment of 8-18 pored scales ending below second dorsal fin, and a posterior segment of 16-28 notched scales, starting 1-2 scale rows below end of the pored segment, and continuing to base of caudal fin. Orbital and anterior nasal cirri simple, the nasal cirrus on the posterior margin of a short tube. Teeth in jaws conical and slightly recurved, a row of larger teeth in front followed by a variable-width band of smaller teeth behind. Vomer always with a single row of conical teeth; palatines edentate or with few small teeth. KEY TO THE WESTERN INDIAN OCEAN SPECIES OF ENNEAPTERYGIUS 1a. Pectoral-fin rays 13-14, all unbranched ............... 2 2a. Pectoral-fin rays 14; belly naked; black spot on margin of second dorsal fin; banded green or brown in life; adults >20 mm SL ........................... E. tutuilae (WIO) 3a. Caudal peduncle with distinct dark bar or marking; body with well defined vertical or oblique dark bars (except E. elegans); anal fin with or without dark bars .. ............................................................................................ 4 4a. Caudal peduncle bar/marking not continuous, but broken in some way ......................................................... 5 5a. Pored lateral-line scales 14-18; anal-fin rays 16 (rarely)-19 .......................................................................... 6 6a. Pored lateral-line scales 16-18 (usually 17); notched lateral-line scales 16-18 (usually 17); anal-fin rays 16-17 (usually 17); caudal peduncle mark two squarish black spots, one above the other; body with indistinct darker bars, red in life; males with lower half of head black . E. elegans (WIO) 7a. Pored lateral-line scales 15-16 (rarely 13-14); anal-fin rays 17-18 .................................................................... 8 8a. Body with 6 distinct vertical dark bars; second dorsal fin with black
spot basally, capped by a bright orange arc in life; caudal peduncle bar broad
with white band at base of caudal fin ........................................................................................... E.
melanospilos (Oman) 9a. Supratemporal sensory canal U-shaped, curving around first 1-2 dorsal-fin
spines ................................ 10 10a. .. Anal fin with 5-7 oblique black bars, basal spots present or absent;
mandibular pores 3+1+3 or 3+2+3 ..11 11a. Anal fin basal spots present; black preanal spot present or absent; body
with broad, diffuse dark bars containing 3-4 pale (white in life) blotches
along mid-side; white line at base of caudal fin; anal-fin rays 19 ...............................................................................
12 12a. .. Pectoral-fin rays 15; second dorsal-fin spines 13; first dorsal fin
not higher than second in males, white in life; caudal peduncle with quadrangular
dusky mark . ............................................................
E. hollemani (Oman) 13a. Pectoral-fin rays 15-16; single symphyseal mandibular pore present; no
labial folds ................... 14 14a. Body with or without scattered melanophores; head of males black or with
cluster of melanophores on cheek.................................................. 15 15a. Males with black head and throat, females with
line from eye onto upper lip in life; body of males with
orange-yellow to yellow-green in life; pored lateral-line
scales 12-15 (usually 13-14) ......................................... 16 16a. Black on head of males to nape, pectoral fin base
and base of pelvic fins; entire body yellow to yellowgreen
in life; females with line of melanophores from eye
onto upper lip; second dorsal fin 11-12 (usually 12); anal
fin rays 17-18 (usually 18)..... ..................... E. abeli (WIO) Enneapterygius abeli (Klausewitz) Tripterygion abeli Klausewitz, 1960: 11, figs. 1-2 (Al
Ghardaqa, Red Sea). DESCRIPTION. Dorsal fins III+XI-XIII+9-10 (usually III+XII+10); anal fin I,17-18 (usually 18); pectoral fins 15: 2-4+4-6+7 (usually 3+5+7). Lateral line with 12-14 pored scales and 19-23 (usually 21- east coast of South Africa - or 22 - Red Sea) notched scales. Notched segment starts one scale row below end of pored segment (Figure 2c); total lateral scales 31; transverse scales 3/6. Vertebrae 10+24-26. Mandibular pores 3+1+3 (Fig. 2d). Head length 3.2-3.8, body depth 4.5-5.2 in SL; eye 2.9-3.7 in head length. Nape scaled, base of first dorsal fin and belly naked. Pelvic fin rays united by membrane for about half length of shorter ray; longest ray reaching mid-vent. First dorsal fin equal in height to second in males, lower in females; second dorsal fin about 60% of body depth. Supratemporal sensory canal crescent-shaped; mouth slightly down-turned, reaching vertical through anterior of pupil; orbital cirrus small and pointed. Live colour. Adult males more darkly pigmented than females or juveniles, with entire head, nape, pre-pectoral area, pelvic fin bases and proximal third of pelvic fins black. Body deep yellow (females) to yellow-green with irregular black markings (males). Females and immature individuals (over 14 mm SL) from Red Sea and Indian Ocean islands with oblique line of melanophores from either side of upper lip to anterior margin of eye, absent in specimens from KwaZulu-Natal, South Africa. Dorsal and caudal fins pinkish with black on margin of first dorsal fin. Clark (1980) records Red Sea male specimens with irregular vertical bars across body, a dark margin to the second dorsal fin and incomplete, slender stripes on third dorsal and caudal fins. Colour in alcohol. Specimens straw-coloured except for black head of males, black margin of first dorsal fin and nasal stripes of females. (See Comparisons below.) DISTRIBUTION. (Fig. 3). Common in Red Sea, along East African coast to southern Kwazulu-Natal, at Mauritius, Seychelles (Randall & van Egmond, 1994), Comoro Islands and St Brandon Shoals; not recorded from Gulf of Oman (Randall, 1995). COMPARISONS. Alive or freshly dead the bright yellow body colour and the black head and chest of the males is distinctive. Without any pigment (females in particular) it is difficult to separate specimens of E. abeli from E. elaine, E. obscurus and E. genamaculatus. In body form and morphometrics the four species are very similar: E. obscurus has higher dorsal fins than E. abeli, the length of the longest spine being sub-equal to body depth, whereas in E. abeli the longest spine is only about 60% of body depth. Enneapterygius abeli may be separated from E. elaine and E. genamaculatus by dorsal- and anal-fin counts. Two of the species also differ in mandibular pore patterns: E. abeli - 3+1+3 and E. obscurus - 2+2+2, while E. abeli and E. elaine have the same pattern. See also under E. elaine. MATERIAL EXAMINED. Red Sea: BPBM 19869 (18.5 mm), Gulf of Aqaba; BPBM 35712 (15 & 16 mm), Hanish Group, southern Red Sea. Comoro Ids: ROM 5470 (12: 11.8-18.6 mm); ROM 5474 (15.3 & 19.1 mm); ROM 67483 (10: 10.3-19.2 mm); ROM 67482 (15: 13.8-18.7 mm); ROM 67480 (17 mm); ROM 67481 (17.5 mm). Mauritius: SAIAB 31508 (19 mm); SAIAB 31511 (18.1 & 18.3 mm); SAIAB 31509 (3: 12.2-17.5 mm); SAIAB 60677 (42: 12.6-23.6 mm); SAIAB 60681 (26:18.5-21.2 mm). St Brandon Shoals: SAIAB 31510 (17.5 mm). Kenya: SAIAB 31506 (9: 17.8-19.2 mm), Shimoni. Mozambique: SAIAB 31507 (3: 16.0-18.5 mm), Ibo Island; SAIAB 31505 (5: 16.5-20.2 mm), Pinda; SAIAB 31513 (19.3 mm), Inhaca Island. South Africa: SAIAB 32418 (67: 10-24 mm); SAIAB 31514 (85:10-23 mm); SAIAB 32415 (33: 9-24 mm); SAIAB 7934 (4: 15.3-21.1 mm); SAIAB 7936 (21.3 & 21.4 mm); SAIAB 9612 (18.3 & 19.5 mm); SAIAB 12362 (17.2 & 17.7mm). Enneapterygius clarkae Holleman Enneapterygius clarkae Holleman, 1982: 121, fig. 6 (Inhaca Island, Mozambique). Enneapterygius n.sp. 2 Clark, 1980: 104, figs. 4a & 14 DESCRIPTION. Dorsal fins III+XI-XII+8-10 (usually III+XII+9); anal fin I,16-17 (usually 16 rays); pectoral fins 15: 2-3+5+6+7 (usually 2+6+7). Lateral line with 11-12 (usually 12) pored scales and 20-22 (usually 22) notched scales, notched segment starting next scale row below end of pored segment (Fig. 4b). Total lateral scales 29 30; transverse scales 3/6. Vertebrae 10+22-23. Mandibular pores 3+2+3 (Fig. 4c). Head length 3.3-3.9, body depth 4.4-5.2 in SL; eye 2.8-3.6 in head length. Nape scaled; belly with thin cycloid scales; single row of cycloid scales on pectoral fin base, parallel with margin of branchiostegal membrane. Pelvic-fin rays united for less than half their length, the longest ray reaching almost to vent. First dorsal fin equal in height to second in females, slightly higher (about 10%) in males. Mouth slightly down-turned and reaches vertical through anterior of pupil. Orbital cirrus length about half pupil diameter, with serrate end; upper and posterior margins of orbits with fine serrations. Supratemporal sensory canal crescent-shaped. Live colour. Body of males and females cream with 45 dark brown transverse bars, which continue across the anal fin, darkest across caudal peduncle, least conspicuous under pectoral fin. Bars may be partially divided, forming Hs or inverted Ys; anal fin bars form basal, subcutaneous black spots; prominent pre-anal spot present. Anterior half of body dusted with melanophores which may form irregular bars on lower portion of head. First dorsal fin dusky; second dorsal fin with irregular dusky bars and one or two black marks on the margin; third dorsal and pectoral fins with irregular dusky markings. Little sexual dichromatism was noted, except that males have red on first and second dorsal fins and are generally more darkly marked than females. Colour in alcohol. Reds and brown fade and specimens retain the bars and spotting as described above. DISTRIBUTION. (Fig. 3) Red Sea, east coast of Africa to northern KwaZulu-Natal, South Africa, the Comoro Islands and St Brandon Shoals; not recorded from Mauritius or Gulf of Oman. COMPARISONS. The species is very similar to E. destai and E. ventermaculus. Fricke (1997) synonymised E. clarkae and E. destai. The bars across the body of E. destai are less well-defined than in E. clarkae, except for the caudal peduncle bar of E. destai which has a distinctive hourglass-shape; that of E. clarkae is solid. The live colours of the two species are also different: bright pink for E. destai and cream and dark brown for E. clarkae. Body bars are absent in E. ventermaculus. Enneapterygius clarkae also has a scaled nape and belly, naked in the other two species. MATERIAL EXAMINED, additional to that listed in Holleman, 1982. Comoro Islands: ROM 67752 (16.5 mm); ROM 67474 (16.4 mm); ROM 67472 (17.0 & 20.5 mm). St Brandon Shoals: USNM 357598 (3: 18.1-19.5 mm); USNM 357609 (20.2 mm). Enneapterygius destai Clark (Fig. 5) Enneapterygius destai Clark, 1980: 102, figs. 4b & 13 (Red Sea). DESCRIPTION (based partly on Clark, 1980; measurements of examined material). Dorsal fins III+XIXIII+8-9 (usually III+XII+8-9); anal fin I,15-17 (usually 16 rays); pectoral fins usually 15: 3,5,7. Lateral line with 8-12 (usually 10-11) pored scales and 21-23 notched scales, notched segment starting second scale row below end of pored segment. Total lateral scales 28; transverse scales 2/5. Vertebrae 10+21-22. Mandibular pores 3+2+3 (Fig. 5b). Head length 3.1-3.3, body depth 4.8-5.1 in SL; eye 2.5-3.0 in head length. Body robust. Scales large, nape scaled, scales to bases of dorsal and anal fins. Two rows of scales at base of caudal fin. Pelvic fins not united by membrane. First dorsal fin lower than second, about 60% of height of second. Pectoral fin of males long, longest ray extending to 2nd or 3rd dorsal-fin ray. Mouth extending to vertical through anterior margin of pupil. Orbital cirrus a rounded flap, about half pupil diameter. Supratemporal sensory canal crescent-shaped. Live colour. Freshly dead adults with head, body and first dorsal fin bright pink, with tints of pink on other fins except pelvics. Five broad oblique dusky bands across body present to some degree in adults, but absent in juveniles. Two prominent black spots on caudal peduncle form an hourglass-shape, upper usually darker than lower. Anal fin with 5, rarely 6, oblique black bars. Prominent black spot present in front of vent on ventral midline. Second and third dorsal fins with irregular dusky bars, second dorsal black in mature males; pectoral fin with 4 vertical dusky bars; 2-3 dusky bars radiate across the lower head from the lower posterior quadrant of the eye. Females generally less intensely coloured than males. Colour in alcohol. Pink fades and the body becomes a pale straw colour, retaining dusky bars on body and fins, the black hour-glass mark across the caudal peduncle and the black bars on the anal fin. DISTRIBUTION. The species is restricted to the Red Sea, where in the south it replaces E. abeli as the most abundant species. COMPARISONS. See under E. clarkae above. MATERIAL EXAMINED. SAIAB 69185 (17.3 mm); SAIAB 69262 (15.0 mm); SAIAB 69204 (4: 15.4-17.3 mm), all from Karaman Island, Uqban,Yemen. Enneapterygius elaine sp.nov. Holotype: SAIAB 65582, male 18.5 mm SL; Rodrigues, Baladirou (ca 19°40' S, 63°26’E); bay with coral rubble; collected P C Heemstra et al., 16 October 2001; sta. ROD-34. Paratypes (all from Rodrigues): AMS 41476-001 (19.6 & 21.0 mm), sta. ROD-37; BPBM 39012 (4: 20.3-21.9 mm), sta. ROD-37; BMNH 2002-8-22:1-3 (3: 17.2-19.2 mm), sta. ROD-34; ROM 73204 (4: 15.3-22.7 mm), sta. ROD-35; SAIAB 65577 (3: 15.0-18.9 mm), sta. ROD-34; SAIAB 65578 (5: 14.6-20.3 mm), sta. ROD-43; SAIAB 65579 (17.4 & 17.6 mm), sta. ROD-04; SAIAB 65580 (3: 17.8-19.1 mm), sta. ROD-27; SAIAB 65581 (3: 21.0-20.7 mm), sta.ROD31; SAIAB 65584 (5: 13.2-21.7 mm), sta. ROD-35; 65586 (5: 15.3-21.7 mm), sta. ROD-37; USNM 370527 (4: 14.120.2), sta. ROD-31. DESCRIPTION. Dorsal fins III + XIII + 10 (rarely with 12 spines and 9-11 rays); anal fin I,17-19; pectoral fins 15: 3+5+7 (rarely 4+4+7); lateral line with 14-15 (usually 14) pored scales and 21-22 notched scales (rarely 20-23), starting second scale row below end of pored segment (Fig. 6b). Total lateral scales 33; transverse scales 2/5. Vertebrae: 10 precaudal, 25 caudal. Mandibular pores 3+1+3 (Fig. 6c). Head 3.2-4.0 in SL; eye 2.4-3.0 in head length. Small fish, seldom longer than 20 mm SL. Body slender, scales large, nape scaled, abdomen naked. Pelvic fin rays slender, united by membrane for half the length of the longer ray. First dorsal fin about half height of second. Pectoral fin large, longest ray reaching to penultimate spine of second dorsal fin. Mouth oblique, small and pointed, maxilla reaching vertical through anterior margin of orbit. Orbital cirrus small and pointed. Live colour. Head of males densely covered with large melanophores, colour dorsally to just behind orbits, laterally to margin of preopercle and ventrally to base of pelvic fins, spotting on lips and throat less dense than elsewhere. Sometimes some brown on lower portion of opercle. Body behind head and including the pectoral-fin base bright yellow, colour extending to junction between first and second dorsal fins and base of pectoral fin rays, giving appearance of a fish wearing a yellow waistcoat. Posterior to this body with light brown reticulations colour generally on margins of scales. Narrow brown bar across caudal peduncle. First dorsal fin pale yellow with a few melanophores on membrane between second and third spines and between third spine and dorsum. Elements of all other median fins light reddish-brown, membranes without colour; caudal fin with narrow yellow-white band at base of rays. Pectoral fin base with two light brown circular spots, one above the other, rays pale brown, membranes immaculate. Head of females forward of hind margin of orbit white with two brown streaks running from eye to upper lip. Head behind eyes brown, colour extending to nape and posterior margin of opercle. Body with same light brown reticulation as males. Median fins as in males, except caudal fin which lacks yellow-white band at base. Pectoral fin bases white, with two light brown circular spots, one above the other, rays pale brown, membranes immaculate. Colour in alcohol. In both males and females all colour fades except for the black of the melanophores on heads of males and on margin of first dorsal fin. ETYMOLOGY. The species is named for Elaine Heemstra, in recognition of her considerable and excellent contribution to the illustration of Indo-Pacific fishes, including several in this paper. The epithet is to be used as a noun in apposition. DISTRIBUTION (Fig. 3). Enneapterygius elaine is currently known only from the lagoon around the island of Rodrigues, western Indian Ocean. COMPARISONS. At first glance Enneapterygius elaine is very similar to E. abeli, but can be easily separated both on colour pattern and counts. The black on the head of male E. abeli extends to the base of the first dorsal fin spine, the posterior margin of the opercles and onto the belly whereas in E. elaine the black extends only to just behind the eyes and to the posterior margin of the preopercle. Enneapterygius elaine usually has 13 spines in the second dorsal fin (usually 12 for E. abeli), usually 19 anal-fin rays (17-18 for E. abeli) and 14-15 pored lateral line scales, compared to a norm of 13 for E. abeli. Female E. elaine also lack the two nasal stripes of preserved female E. abeli specimens. Enneapterygius elegans (Peters) Tripterygium elegans Peters, 1876: 441 (Mauritius) (lectotype ZMB 9465, designated by Fricke, 1997: 182). Enneapterygius elegans: Holleman, 1986: 756, pl. 116, figure 236.4 ; Fricke & Randall, 1992: 3 (Maldives);. Fricke, 1994: 203. DESCRIPTION. Dorsal fins III+XI-XIII+8-10 (usually III+XII+9); anal fin I,16-17 (usually 17 rays); pectoral fins 16: 2-3+6-7+7. Lateral line with 16-18 pored scales and 16-18 notched scales, notched segment starting one scale row below end of pored segment, overlapping by 2-3 scales (Figure 7c).Total lateral scaled 30-31; transverse scales 3/5. Mandibular pores 6-9+1+6-9 (Figure 7d). Head length 3.2-3.6, body depth 3.8-4.6 in SL; eye 3.0 3.7 in head length. Body robust. Nape scaled; belly with thin cycloid scales, entire in males, posterior half only in females. Pelvic-fin rays united by membrane for half length of the shorter ray, longer ray extending three-quarters distance to vent. First dorsal fin about two-thirds height of second dorsal fin, height of second half to two-thirds body depth. Mouth slightly oblique, extending to vertical through anterior margin of pupil. Orbital cirrus small and simple. Exposed posterior margins of median and lateral extrascapulars finely serrate. Supratemporal sensory canal crescent-shaped. Live colour. Males with brick-red body and lighter, irregular, narrow bars. Head from below middle of eye, including upper lip, densely spotted with melanophores, extending onto throat, pectoral fin bases and anterior of belly; nasal area above upper lip not spotted. Caudal peduncle distinctly marked with two squarish, black spots separated by a mid-lateral unpigmented space, often giving the impression of a dumbbell or hourglass; bright yellow spot on the peduncle just posterior to the last dorsal-fin ray. First and second dorsal fins stippled with red and black, second dorsal darker along margin and base, third dorsal with broken orangey bars and black stipples. Caudal fin with a narrow red stripe at its base; anal fin red; pectoral fins red with creamy blotch at base of central rays; pelvics stippled with red. Females less intensely marked: body with indistinct bands with light spots between; no black on lower part of head and throat and fewer melanophores on head and fins. Belly and pelvic fins white; stipples along the base and margin of second dorsal fin indistinct. Colour in alcohol. All colour fades and specimens are pale brown, sometimes with indistinct banding on body and always with twin black blotches on caudal peduncle and black on head and throat of males. DISTRIBUTION (Fig. 3). Mauritius, Seychelles, Comoro Islands, St Brandon Shoals and Maldives as well as along the east coast of Africa from Kenya to Bazaruto, Mozambique. It is also widely distributed through the Indo-West Pacific Ocean (see Fricke, 1994). COMPARISONS. Small Enneapterygius elegans may be confused with specimens of E. destai or E. gruschkai, both of which have broken bars across the caudal peduncle, that of E. destai also two spots, one above the other. Enneapterygius destai is however confined to the Red Sea. Enneapterygius elegans can be readily separated from E. gruschkai, with which it occurs sympatrically in the western Indian Ocean, by lateral line count: usually 17+17 for E. elegans vs. usually 15+20-21 for E. gruschkai. MATERIAL EXAMINED. Mauritius: BPBM 20171 (25 & 27 mm); SAIAB 61342 (19: 17.9-25.4 mm); SAIAB 60614 (4: 10.5-20.8 mm). Seychelles: SAIAB 7710 (10: 14.3-27.3 mm), Mahé; SAIAB 950 (41: 19.5-29.5 mm), La Digue. Comoro Ids: ROM 67466 (4: 16.3-20.5 mm). St Brandon Shoals: USNM 218483 (27.0 & 29.7 mm); USNM 218764 (4); USNM 218765 (26.5 & 29.7 mm); USNM 218766 (22: 18-23.8 mm); USNM 218767 (17: 19.4-30.0 mm). Kenya: SAIAB 951 (6: 19.2-20.7 mm), Shimoni. Mozambique: SAIAB 7700 (26.1 mm), Pinda; SAIAB 7702 (23.5 & 28.5 mm), Pinda; SAIAB 7699 (4:13.8-28.2 mm), Ibo; SAIAB 7698 (27.4 & 28.3 mm), Bazaruto. Enneapterygius fasciatus (Weber) (Fig. 8) Tripterygium fasciatum Weber, 1909: 148 (Savu [Sawu], southern Indonesia and the Karakelang [Karakelong] Islands, northern Indonesia). Tripterygion (Enneapterygius) fasciatum: Mukerji, 1935: 272 Tripterygion fasciatum: Herre, 1939: 351 Tripterygion fasciatus: Lal Mohan, 1968: 116, figure 2 DESCRIPTION. Dorsal fins III+XI-XIII+9-11 (usually III+XII+9-10); anal fin I,17; pectoral fins15-16: 3-4+4-5+78. Lateral line with 13-16 (usually 15-16) pored and 1821 notched scales, notched segment starting one scale row below end of pored segment and overlapping it by 2 -3 scales (Fig. 8b). Total lateral scales 30-32; transverse scales 3/6. Mandibular pores 3+1+3 (Figure 8c). Head length 3.3-3.5, body depth 3.9-4.6 in SL; eye 2.9-3.2 in head length. Body robust. Nape scaled, belly naked. Pelvic fins united by membrane for two-thirds of length of shorter ray, longest ray reaching anus. First dorsal fin lower than second, second about 70% of body depth. Supratemporal sensory canal crescent-shaped. Mouth reaching vertical through anterior margin of pupil; orbital cirrus small and lobate. Live colour. Weber’s original description of the species gives live colours of a yellowish body with seven brown crossbars, head with brown flecks and bands, a dark base to the first and second dorsal fins and seven bar-like flecks on the anal fin. Herre (1939: 397) gives a colour description that closely agrees with that of Weber. A more recent description by Lal Mohan (1968: 117), based on a single specimen, reads: “Cheeks, snout, anterior part of belly mottled with blue chromatophores, a dark blotch on first dorsal between first and second spines; five vertical irregular greenish bands on body, a brown blotch on lower part of opercle, a reddish blotch on base of pectoral peduncle and ventral fin, six deep blue spots on base of anal fin.” Herre however noted a fairly distinctive dark band at the base of the caudal fin not mentioned by Lal Mohan. His illustration, as does that of Herre, shows the body bars as oblique, which is in agreement with the specimens I have seen. The 6-8 black (blue) spots at the base of the anal fin of both sexes are noteworthy. Colour in alcohol. Body of males light brown with faint bands of melanophores and 6-8 distinctive basal anal-fin spots. Even scatter of melanophores cover head to pectoral fin bases and anterior portion of belly. Dorsal and anal fins with light scatter of melanophores. Females lack the dark head, show more obvious banding and also carry the basal anal fin spots. DISTRIBUTION (Fig. 3). East coast of Africa, the Seychelles, the southeast coast of India, Sri Lanka, the Andaman Islands and as far east as the Solomon Islands (Fricke, 1997: 195). COMPARISONS. Enneapterygius fasciatus is similar in size and robustness to E. elegans, but lacks the scaled abdomen and conspicuous peduncular mark of that species (note: not the band at the base of the caudal fin). Their live colours also separate them: greenish with brown banding for E. fasciatus and brick-red for E .elegans. MATERIAL EXAMINED. Kenya: SAIAB 60612 (7: 17.5-22.2 mm), Malindi; SAIAB 60682 (23.0 mm), Shimoni; LACM 31617-27 (25.8 mm), Manda Island. Tanzania: SAIAB 60683 (28.3 & 29.5 mm), Zanzibar. Mozambique: SAIAB 60609 (25.3 mm), Ibo; SAIAB 60610 (28.1 mm) Mocimboa da Praia. Comoro Ids: ROM 5474 (15.3 & 20.3 mm). Enneapterygius genamaculatus sp. nov. (Fig. 10) Holotype : USNM 357600, male 22.3 mm SL; St Brandon Shoals, SE side of Grande Passe (ca 16°28' S, 59°40' E); dead and rocky reef channels and ledges; collected V G Springer et al. 5 April 1976; sta.VGS 76-9. Paratypes (all St Brandon Shoals): SAIAB 60503 (6: 19.321.4 mm), same collection as USNM 357602; USNM 357597 (17.8 & 21.0 mm), sta.VGS 76-22; USNM 357606 (17.5 & 19.7 mm), sta. VGS 76-7; USNM 357602 (12: 19.522.8 mm), sta.VGS 76-10; USNM 357599 (29: 17.6-22.9 mm), sta.VGS 76-9. DESCRIPTION. Dorsal fins III+XII-XIII+9-11 (usually III+XIII+10); anal fin I,18-19 (usually 19 rays); pectoral fins 15-16: 3-4+5-6+7-8. Lateral line with 13-15 pored and 21-23 notched scales, notched segment starting one scale row below end of pored segment (Fig. 10b). Total lateral scales 32-33; transverse scales 2/4. Vertebrae 10+24. Mandibular pores 3+1+3 (Fig. 10c). Head length 3.5-3.9, body depth 4.6-5.4 in SL; eye 2.7-3.1 in head length. Small (less than 25 mm SL), slender fishes; nape scaled, belly naked. Pelvic-fin rays united by membrane for entire length of shorter ray, longer ray reaching anal-fin spine. First dorsal fin lower than second; maximum height of second dorsal fin about 70% of body depth. Supratemporal sensory canal crescent-shaped. Orbital cirrus small and rounded. Tip of tongue narrow and rounded; palatines with teeth; maxilla reaching vertical through anterior margin of pupil. Live colour. The live colour of the species is not known. Colour in alcohol. Specimens straw-coloured. Males with large cluster of melanophores below and slightly posterior to the eye. Large specimens may have some pigment on the snout, in the interorbital area, on top of the head and on the membrane across the throat. Two small clusters of melanophores at upper and lower ends of pectoral-fin base and small black spots on membrane between third dorsal-fin spine and dorsum. Females with only the two clusters of melanophores at the corners of the pectoral-fin base and a few melanophores on the first dorsal-fin membrane. DISTRIBUTION (Fig. 9). The only known specimens are those collected by Springer et al. from St Brandon Shoals. ETYMOLOGY. The name is taken from the Latin gena, a cheek, and macula, a spot, and is given for the spots on the cheeks of the males of the species. COMPARISONS. Morphometrically the species is similar to E. gruschkai, but lacks the body markings of that species. Unmarked specimens would be impossible to separate except on mandibular pore patterns: 3+1+3 for E. genamaculatus and 3+2+3 for E. gruschkai. Enneapterygius gruschkai sp.nov. Holotype: SAIAB 61281; male, 24.3 mm SL; Passe dAmbulante, Mauritius (20°26’10"S, 57°17’40"E); depth 6-8m, spur and groove formation; collected P. C. Heemstra et al., 18 May 1995; sta. PCH 95-M30. Paratypes: Seychelles: BPBM 35600 (28.1 mm). Comoro Ids: CAS 33424 (23.3 mm); ROM 71834 (12.1 & 23.7 mm); ROM 71833 (10: 10.4-25.2 mm). Chagos Archipelago: ROM 56795 (4: 17.8-23.1 mm); ROM 56796 (18.4 mm); ROM 56797 (21.8 mm); ROM 56798 (23.2 mm); ROM 56799 (20.5 & 22.0 mm); ROM 56800 (20.0 mm); ROM 56801 (5: 16.6-24.4 mm); ROM 56802 (6: 17.3-25.2 mm); USNM 357612 (25.8 mm). St Brandon Shoals: USNM 357601 (6: 22.6-29.2 mm); USNM 357607 (23.7 mm); USNM 357611 (22.3 & 24.5 mm); USNM 357604 (22.7 & 24.1 mm); USNM 357596 (5: 23.0-24.7 mm); USNM 357608 (8: 23.2-27.0 mm). Mauritius: SAIAB 61282 (11: 13.0-26.3 mm); SAIAB 61283 (11: 22.4-25.2 mm). Non-types: Comoro Ids: CAS 34507 (21.3 mm). St Brandon Shoals: USNM (VGS 76-18) (22.1 mm). DESCRIPTION. Dorsal fin III+XII-XIII+8-11 (usually III+XIII+9-10); anal fin I,17-19 (usually 18-19 rays); pectoral fins 15: 2+7+6 (occasionally 3+6+6); lateral line with 14-16 (usually 15) pored scales and 19-23 (usually 20-21) notched scales, notched segment starting next scale row below end of pored segment, below mid-second dorsal fin (Fig. 11b). Total lateral scales 33; transverse scales 3/5. Vertebrae 10+25. Mandibular pores 3+2+3 (Fig. 11c). Head length 3.1-3.6, body depth 5.4-5.9 in SL; eye 2.7-3.2 in head length. A medium-sized species of Enneapterygius (less than 30 mm SL). Nape scaled, body scales continuing to bases of dorsal and anal fins; belly naked. Pelvic-fin rays not united by membrane. First dorsal fin equal in height to second, second about 70% of body depth. Simple, lobate orbital and anterior nasal cirri present. Supratemporal sensory canal slightly curved. Tongue triangular and sharply pointed; maxilla reaches vertical through anterior margin of pupil; broad band of teeth in upper and front of lower jaw; palatines without teeth. Spines of first dorsal fin with small, laterally projecting spinelets. Live colour. Colour of freshly caught males varies from 5 dark brown bars on creamy-yellow ground body colour, with bars sometimes divided below midline, to a rich brown body with 5 narrow creamy-white bars, the last at the base of the caudal fin. Darkest bar across caudal peduncle with small, clear window mid-laterally; peduncle cream or yellow either side of bar. Penultimate two bars darkest dorsally, forming a small dark saddles at base of last two dorsal-fin rays and on caudal peduncle. Scale-pocket margins with brown and orange spots. Head markings dark brown to black, the face almost tiger-striped; top of head deep orange with black stippling. First dorsal fin with black stipples; second with orange bands on spines and black stipples on membranes; third same as second, but paler. Anal fin stippled black except for tips of rays, which are colourless. Pectoral fins with a black spot near dorsal and ventral ends of base, a white blotch posterior to the lower black spot, followed by another black spot and a larger white blotch in the centre of the fin; rays orange distally. Caudal fin deep orange to red proximally, with narrow band of black stipples at its base, and dusky distally. Females and juveniles less heavily pigmented, body bars and facial markings pale brown; dorsal fins with yellow-orange pigment only and no black stippling; anal fin pale yellow with some black stippling; caudal fin with orange and black stippling at base of rays. Colour in alcohol. All colour except black fades. Large males with 5 dark bars across pale cream body, darkest across caudal peduncle, with characteristic window at centre, and dark saddle at base of last two dorsal-fin rays. Face tiger-striped. Females and juveniles may lose all colour except for the peduncular bar and saddle. Juveniles, females and less heavily pigmented males may have no markings except for the double saddle mark on the caudal peduncle. ETYMOLOGY. The species is named for Victor Gruschka Springer, doyen of blennioid systematics. DISTRIBUTION (Fig. 9). The species seems to be restricted to the islands of the western and central Indian Ocean and has not been recorded from the east coast of Africa. COMPARISONS. Enneapterygius gruschkai may be mistaken for small specimens of E. elegans, but the former species has 18-19 anal-fin rays compared to 16-17 for E. elegans and 15 pored and 20-21 notched lateral-line scales compared to 16-18+16-18 for E. elegans. Enneapterygius gruschkai also lacks the scaled belly of E. elegans. Enneapterygius hollemani Randall Enneapterygius hollemani Randall, 1995: 27, figure 1 (Gulf of Oman). DESCRIPTION (from Randall, 1995). Dorsal fins III+XIII+10; anal fin I,19; pectoral fins 15: 2-3+6+6-7. Lateral line with 16 pored and 23-24, notched scales starting second scale row below end of pored segment and overlapping it by a single scale. Mandibular pores 3+1+3. Head length 3.3 in SL; eye 3.2-3.4 in head length; body depth 4.6-5.2 in SL. Body moderately elongate. Nape scaled except adjacent to anterior of first dorsal fin, belly naked. Pelvic-fin rays united by membrane for half of length of shorter ray, longest ray reaching anus. First dorsal fin equal in height to second, about 65% of body depth. Supratemporal sensory canal U-shaped. Mouth small, reaching vertical through anterior of orbit. Orbital cirrus very small and lobate. Live colour. Body of males pale green, edges of scales with red and dark brown spots forming 3 broad dusky bars which divide ventrally, 2 below second dorsal fin and one below third dorsal fin. Bars darker ventrally than dorsally and separated by clusters of white flecks. Quadrangular dark mark on caudal peduncle, its rear margin a narrow dark brown bar at base of caudal fin. Belly white, anus edged in black; black preanal spot may or may not be present. Head a mixture of pale green, red and brown, becoming pale green ventrally. Upper lip dusky; orbital cirrus white. First dorsal fin creamy-white with red tip to first spine; second dorsal fin with red cross-bands on spines, 3 oblique white bands and a dusky band at its base; third dorsal fin similar but paler. Anal fin with 7 black spots basally, which continue internally and diagonally across fin with white in between; rays with white and dark red spots. Pectoral fin rays white with red edges and 6 dark brown spots forming 6 irregular transverse bars, lower rays orange-yellow distally; membranes pale. Pelvic fins white. Caudal fin with narrow white band at base, transparent membranes and bright green rays. Colour in alcohol. Body tan with melanophores along scale margins except ventrally, forming bands as described above. Quadrangular dusky mark on peduncle with narrow dark band at base of caudal fin; head below eyes dusky, cheeks paler. Small dark brown spot just anterior to origin of lateral line and a second spot of same size just postero-ventral to the first. Anus edged in black, black preanal spot present in 2 of the 3 specimens on which the original description is based. First dorsal fin slightly dusky; second dorsal fin with broad dusky band at base; third dorsal and caudal fin pale. Anal fin with 7 black basal spots which continue subcutaneously and as diagonal bands across the fin. Prepectoral region dusky, darkest ventrally at base of fin, continuing as a pointed dark blotch below branchiostegal membranes. Pectoral fins pale with dark brown spots on rays forming 6 irregular transverse bands. DISTRIBUTION (Fig. 9). Described from the Gulf of Oman where 3 specimens were collected from shallow tidepools along a rocky shore. COMPARISONS. Enneapterygius hollemani is similar in colour to E. ventermaculus, but differs in the shape of the peduncular mark (quadrangular in E. hollemani, triangular in E. ventermaculus), the colour of the first dorsal fin (brown and orange in E. ventermaculus; white in E. hollemani), and pelvic fins (yellowish in E. ventermaculus, white in E. hollemani) and on the head (bottle green with more red and orange in E. ventermaculus, greenish in E. hollemani). Enneapterygius hollemani also has 15 pectoral fin rays vs. 13 (rarely) or 14 for E. ventermaculus; 16 pored lateral line scales vs. a mode of 15 for E. ventermaculus and 13 spines in the second dorsal fin vs. a mode of 12 for E. ventermaculus. Other differences are detailed in Randall (1995: 29). Enneapterygius kosiensis sp. nov. (Fig. 12) Holotype. SAIAB 59086 (19.4 mm SL), male, Sodwana Bay, KwaZulu-Natal, South Africa, coll. P. C. Heemstra et al. 10 August 1998; sta. FW 98-12. Paratypes. SAIAB, 40265 (9: 14.5-17.1 mm SL ), reef 2 km south of Kosi Mouth, KwaZulu-Natal, South Africa, coll. C. D. Buxton et al., 9 August 1992; sta. 1-92-11. SAIAB 9897 (18.1 mm SL) Sodwana Bay, KwaZulu-Natal, coll. P. C. Heemstra et al., 22 April 1979; sta. PCH 79-27. DESCRIPTION. Dorsal fins III+XII+9; anal fin I,16-17 (usually 17 rays); pectoral fins 13, all rays simple; lateral line with 12 pored scales and 21-22 notched scales, notched segment starting second scale row below end of pored segment, overlapping by one scale (Fig. 12b). Total lateral scales 32; transverse scales 1/4. Vertebrae 10+22. Mandibular pores 2+1/2+2 (Fig. 12c). Head 3.2-3.5, body depth 5.1-5.3 in SL; eye 3.1-3.7 in head length; maxilla 2.6-2.9 in head length. Small fish, less than 20 mm SL. Body slender; scales large; nape scaled; belly with smaller, thin cycloid scales. Pelvic fins slender, rays united by a thin, fragile membrane for about one third length of shorter ray. First dorsal fin slightly higher than second, second 75-90% of body depth. Longest pectoral-fin ray reaches nearly to base of first ray of third dorsal fin. Supratemporal sensory canal crescent-shaped. Mouth oblique, reaching vertical through anterior margin of pupil; interorbital concave and broad, about equal to pupil diameter; orbital cirrus a simple rounded flap and about half pupil diameter in length. Live colour. Not known. Colour in alcohol. Entire fish a pale straw colour. Males with a few melanophores scattered on body and a cluster of large melanophores on midside, beneath pectoral fin, and a cluster of melanophores on the caudal peduncle below the midline. Margin of first dorsal fin with cluster of large melanophores, a band of melanophores parallel to margin of second dorsal fin and 6 irregular black bars on anal fin. Basal spots may be present subcutaneously on anal fin and a preanal spot may be present. Females lack the fin markings and have fewer melanophores on the body. ETYMOLOGY. The species in named after Kosi Bay, the area off which specimens were first found. DISTRIBUTION (Fig. 9). Currently known only from northern KwaZulu-Natal, South Africa (Sodwana Bay and Kosi Bay), taken on flat reef at 27-30m depth. COMPARISONS. Preserved specimens of Enneapterygius kosiensis may be mistaken for small specimens of E. ventermaculus; E. kosiensis, however, has a scaled belly and 2+1/2+2 mandibular pores, compared to 3+1+3 for E. ventermaculus and 13 simple pectoral-fin rays (vs. 14 for E. ventermaculus, of which 4-6 are divided). Only one other western Indian Ocean species, E.tutuilae, has all pectoral fin rays simple, 14 in number. Enneapterygius melanospilos Randall (Plate 1) Enneapterygius melanospilos Randall, 1995: 29, fig. 2 (Arabian Sea, central coast of Oman). DESCRIPTION. (from Randall, 1995). Dorsal fins III+XII+10; anal fin I,18; pectoral fins 15-16: 2+5-6+8; lateral line with 14-15 pored and 21-22 notched scales, notched segment starting one scale row below end of pored segment. Total lateral scales 34. Mandibular pores 3+2+3. Head 3.2 in SL; eye 2.7-3.0 in head length; depth 4.8-5.5 in SL. Body moderately elongate. Nape scaled, belly naked. Pelvic-fin rays scarcely united by membrane. First dorsal fin higher than second in males (by about 20%) and about the same height as second in females, at about 75% of body depth. Numerous small spinules along posterior edge of supratemporal sensory canal and on upper posterior edge of orbit. Supratemporal sensory canal crescent-shaped. Mouth not large, reaching vertical though anterior of orbit. Orbital cirrus bilobed or trilobed, equal in length to pupil diameter. Live colour. Body whitish with 6 orange-brown bars, becoming progressively darker posteriorly, the darkest across the caudal peduncle. Width of last 4 bars about equal to white interspaces. Front, top of head and nape brown mottled with orange; lower half of head pale with brown-edged orange stripe below eye. Opercle dark orangey-brown shading to yellowish posteriorly. First dorsal fin pinkish; second with jet-black spot as large as eye basally between 7th and 11th spines, fin white above spot with bright orange arc on margin, remainder of fin transparent with irregular oblique dusky bars with orange on spines; third dorsal fin with 2 brownish-orange bands, continuous with 4th and 5th body bars; anal fin whitish with 4 broad dusky zones; caudal fin with white band at base, membranes clear and rays pink, becoming whitish at tips; pectoral fins with clear membranes, upper rays pale yellowish, lower rays yellow; pelvic fins white. Colour in alcohol. Orange and yellow colours fade and body becomes whitish with 6 dusky bands, head mottled, stripe below eye, distinctive mark on second dorsal fin and banding on other fins. DISTRIBUTION (Fig. 9). Described from three specimens taken in the Arabian Sea off the coast of Oman. COMPARISONS. Enneapterygius melanospilos shares the distinctive banded pattern on body with E. clarkae and E. gruschkai and is similar in colour to E. gruschkai, which also has an orange and brown colour pattern. The spot on the second dorsal fin distinguishes E. melanospilos from the other two species. Randall also notes the long bi- or trilobed orbital cirrus, the spinules on the upper posterior rim of the eye and along the posterior margin of the supratemporal sensory canal as distinctive. The long orbital cirrus is distinctive, but the serrate margin of the median extrascapulars is also found in E. elegans and the spinules on the rim of the eye in E. clarkae. Enneapterygius clarkae has fewer pored lateral-line scales than E. melanospilos (12 vs. 14-15) and E. gruschkai generally has 13 spines in the second dorsal fin (vs. 12 for E. melanospilos) and the distinctive window in the bar across the caudal peduncle. Enneapterygius obscurus Clark Enneapterygius obscurus Clark, 1980: 105, figs. 4d & 15 (Aqaba & Eritrea, Red Sea); Fricke & Randall, 1992: 6, fig. 3 (Maldives). DESCRIPTION (partly from Clark 1980). Dorsal fins III+XII-XIV+9-10, usually III+XIII+9-10; anal fin I,16-18 (usually 17-18 rays); pectoral fins 14-15: 3-4+5+6; lateral line with 10-12 pored and 21-22 notched scales, pored segment starting 2 scale rows below end of pored segment (Fig. 13b). (In some of the Red Sea types examined there were only 7-8 pored scales, followed by 2 notched scales - Fig. 13c.) Transverse scales 1/4. Mandibular pores 2+2+2 (Fig. 13d). Head 3.2-4.0, body depth 6.5 in SL; eye 2.4-2.6 in head length. Body moderately slender. Nape scaled, belly naked. First dorsal fin equal to or a little lower than second, second subequal to body depth. Pelvic-fin rays united by membrane for half length of shorter ray. Profile rounded; mouth low on head, cleft nearly horizontal, reaching vertical through anterior margin of pupil. Prominent labial folds extend from lower lip, under the jaw, in a crescent to the mandibular sensory canal (Fig. 13d). Supratemporal sensory canal crescent-shaped. Orbital cirrus bilobed and moderately long, about half pupil diameter. Live colour. There are three transparencies, one taken by Randall in the southern Red Sea and two in Debelius’ Red Sea Reef Guide (1998: 174). The Randall specimen was captured and identified as E. obscurus, while the specimens illustrated in Debelius were not collected or identified. The fin element counts that can be made from the Debelius illustrations agree with those of E. obscurus and not with those of E. pallidus, another Red Sea species for which the live colour is unknown. The Debelius illustrations also show the irregular bands of melanophores on the second dorsal fin, the black-edged scales above the lateral line, between the second and third dorsal fins and a small, black spot at the upper base of the caudal peduncle, which shows as deep red. While the cluster of black spots on the lower base of the pectoral fins is not visible, the other features confirm identification of the Debelius illustrations as those of E. obscurus. Body of males transparent with irregular crimson markings, scale margins lined with crimson; large crimson blotch below pectoral fin, and two small deep crimson spots at top and bottom of the caudal peduncle. Vertical line of scales above lateral line, between the second and third dorsal fins, edged in black. Opercles crimson; upper portion of head crimson with some white marks; eye crimson, except for a fine yellow line around the pupil. Orbital cirri yellow. First dorsal fin deep crimson anteriorly with yellow markings, white posteriorly with black spots. Elements of second and third dorsal fins crimson with irregular bands of melanophores on membranes. Anal, caudal and pectoral-fin elements pale red, membranes transparent. (Preserved specimens show melanophores on the membranes of the anal fin, banded in females.) Females are lighter in colour, with pale red, irregular bands on the body, and lack the intense red on the head and brightly coloured first dorsal fin of males. Colour in alcohol. Specimens pale with no conspicuous markings except a cluster of few melanophores on lower pectoral-fin base, a small black spot on upper caudal peduncle and a vertical line of a few black-edged scales on the body below the end of the second dorsal fin. Few melanophores on head, just above anterior part of pterotic sensory canal. Second and third dorsal fins with irregular bands of melanophores; anal fin with melanophores evenly scattered on membranes in males, banded in females. Dorsal and anal fins of females with weak oblique, dusky bands. Pectoral fins with scattered melanophores on mid- and upper-base and on membranes of proximal two-thirds of fin. DISTRIBUTION (Figs. 9, 17). Enneapterygius obscurus was originally described from the Red Sea (Aqaba and Eritrea) and was presumed endemic to the area. Randall (1995) does not record the species from Oman, where the species may reasonably be expected to occur if it does occur outside the Red Sea. Fricke & Randall (1992) do, however, record a single specimen from the Maldives which agrees morphometrically with Clark’s (1980) description, but differs considerably in pigmentation, having 6 narrow bars across the body. Randall collected a specimen in the Seychelles that has similar pigmentation. I have seen a single specimen from Malindi, Kenya that can only be identified as E. obscurus, except that it has a single symphyseal pore. It is then possible that the species is more widespread in the western Indian Ocean. COMPARISONS. There seems little to distinguish preserved material of E. obscurus from that of several other species, other that the lack of any obvious pigmentation except the cluster of melanophores at the pectoral-fin base and the spot on the upper caudal peduncle, the height of the first dorsal fin and shape of the supratemporal sensory canal. Morphometrically E. obscurus is similar to E. abeli, E. clarkae, E. destai and E. ventermaculus, except that E. obscurus is the only species in this group with a 2+2+2 mandibular pore pattern. The other three species usually have distinguishing pigmentation, but female E. abeli specimens do not. The height of the first dorsal fin of E. abeli is only about half body depth compared to being subequal to body depth in E. obscurus. The scarlet live colour of the species is, however, very distinctive. MATERIAL EXAMINED. Red Sea: BPBM 35721 (17.8 mm), southern Red Sea; HUJ 5486 (4: 14.5-16.2 mm), paratypes, El Himera; HUJ 5483 (17.5 & 22.0 mm), paratypes, El Himera. East Africa: SAIAB 30422 (24.3 mm), Malindi, Kenya. Enneapterygius pallidus Clark Enneapterygius pallidus Clark, 1980: 107, Figures 4e & 16 (Aqaba & Eritrea, Red Sea). DESCRIPTION (from Clark 1980). Dorsal fin III+XIIIXIV+10-11; anal fin I, 21-22; pectoral fins 13-14: 3+56+5. Lateral line with 10-11 pored scales and 28(?) notched scales, notched segment starting second scale row below end of pored segment. Mandibular pores 2+2+2; lower jaw with two large lip flaps (Figure 14b). Head 3.4-4.7, body depth 6.9 in SL. Body very slender. Nape and belly naked. First dorsal fin equal to or lower than second, second about equal to body depth. Pelvic-fin rays united by membrane for about half of length of shorter ray. Supratemporal sensory canal deeply U-shaped. Orbital cirrus small and lobate. Live colour. Recent underwater photographs taken by Richard Field at Farasan Island in the southern Red Sea of an unidentified tripterygiid may possibly be of E. pallidus. With the identification of Debelius’ photographs as E. obscurus (see above) and Randall’s photographs, E. pallidus remains the only Red Sea species for which the live colour is unknown. Thus, unless Field’s photograph represents an undescribed species, by a process of elimination, it is of E. pallidus. Body translucent with faint red bars, the most prominent across the caudal peduncle. Top of head opercle and anterior of body bright green; iris green and brown; fins translucent with a few dark spots on membrane between first two dorsal-fin spines. The apparent absence of melanophores suggests that this is E. pallidus, but will have to be confirmed. Colour in alcohol. No sexual dichromatism evident. There are no distinguishing marks. Few melanophores over the brain area, a few on mid-opercle and a few on pectoral-fin base. DISTRIBUTION (Fig. 15). The species was described from four specimens from the Red Sea (Aqaba and Eritrea). COMPARISONS. Fricke (1997) considers the species synonymous with E. pusillus. It can however be distinguished from all WIO tripterygiids by its high anal-fin ray count (21-22 rays), and that only the lower 5 pectoral rays are simple (for E. destai as well), compared to 6-8 for other species. Clark (1980) also records that E. pallidus has 11+26-28 vertebrae compared to 11-12+2225 for E. pusillus. Enneapterygius pusillus is further distinguished from E. pallidus by the former’s very high first dorsal fin. Enneapterygius philippinus (Peters) Tripterygium philippinum Peters, 1868: 269 (Luzon Island, Philippines). Tripterygium minutum: Günther 1877: 211(Samoa). Enneapterygius tusitalae: Jordan & Seale 1906: 416, fig. 97 (Samoa). Enneapterygius pardochir: Jordan & Seale 1906: 417, fig. 98 (Samoa). Enneapterygius punctulatus: Herre 1935: 432 (New Hebrides). (See Fricke 1997: 274 & 276 for a detailed synonymy). DESCRIPTION (based partially on Fricke, 1994 & 1997; note that western Pacific Ocean specimens exhibit a wider range of counts than specimens from the western Indian Ocean - see Fricke 1994: 249; 1997: 287 & 288): Dorsal fins III+X-XIII+8-9 (usually III+XI+9); anal fin I,1517, (usually 16 rays); pectoral fins 15: 3-4+4-5+7; lateral line with 10-15 (usually 12-13) pored scales and 20-22 notched scales, notched segment starting second scale row below end of pored segment (Figure 15c). Total lateral scales 29-31; transverse scales 2/5. Mandibular pores 23+1+2-3 (Fig. 15d). Head 3.6-4.2, body depth 5.0-5.6 in SL; eye 2.8-3.2 in head length. Small, slender fishes, less than 22 mm SL. Nape, belly and pectoral-fin bases naked. Pelvic-fin rays slender and united by membrane for two-thirds of length of shorter ray. First dorsal fin lower than second. Mouth slightly oblique, reaching vertical through anterior margin of orbit. Orbital cirrus small and lobate. Supratemporal sensory canal crescent-shaped. Live colour. Fricke (1994: 245) quotes a description by J. B. Hutchins of the colour of specimens after two weeks in formalin: “Upper sides green-yellow, lower sides red-yellow. All scales on sides profusely spotted in black, also red spots on dorsal surface. Lower half of head with larger black spots extending onto pectoral [fin base]. Breast and throat pale with black spots. Eye red. Caudal, dorsal and anal fins spotted with black.” Jordan and Seale’s (1906) description of E. tusitalae is in reasonable agreement with Hutchins, but note that E. pardochir (synonymised with E. minutus by Fricke, 1994) varies from, “cherry red with darker bars, most distinct and paired alongside, iris scarlet, fins all reddish, barred with brown...” to “...bright green with bronze dots, sides with large white spots, two white dots at base of caudal, pectoral banded white with reddish brown...” Underwater photographs of a male and a female in Masuda & Kobayashi (1994: 310) show the following: male with body, head and fins liberally covered with densely packed melanophores with some white flecks on body, first and second dorsal fin and head. Females with less black, showing clearer banding on body and fins. Photographs of a male and female by Winterbottom of fresh specimens from Lizard Island, Australia, agree with Hutchins description and a photograph by Heemstra of a single male from Rodrigues (Plate 2) shows the top of the head and anterior dorsum of the body as dark yellowish-green. Colour in alcohol. Body of mature males with melanophores evenly and densely spread over body and fins, the larger ones on cheeks, thorax and along the dorsum, black in the centre surrounded by a brown halo. Lower half of head darker and in large males becomes uniformly brown-black. All fins densely covered with melanophores; dorsal fins darker distally; anal, caudal and pectoral fins very dark, particularly lower half of caudal and pectorals, the latter with melanophores on inner surface as well. Immature males and females paler, with 4 oblique black bars across the body, with white blotches between, the bars dividing ventrally to continue as narrower bars across the anal fin. First and second dorsal fins evenly dark, pigment on spines; third dorsal without colour; caudal and pectoral fins irregularly barred. Females have no pigment on throat, belly or along the base of the anal fin. DISTRIBUTION (Fig. 9). The species is widely distributed throughout the Indo-West Pacific Ocean. It was originally described from the Philippines (Peters, 1868), Günther (1877) described the species from Samoa; Herre (1935) from the New Hebrides; Schultz (1960) from the Marshall Islands; Fowler (1958) from the Ryu Kyu Islands, and Winterbottom collected material at Lizard Island (Australia) in 1981. In the Indian Ocean the species has been recorded from St Brandon Shoals, the Comoro Islands, and several island localities along the East African coast. There is a single collection of 16 specimens from the Great Fish River Point in the Eastern Cape, South Africa, made by J. L. B. and M. M. Smith in 1954, but the locality is highly doubtful. COMPARISONS. Male Enneapterygius philippinus, with their distinctive dense covering of melanophores, are easily distinguished from other species of the genus. Females, sometimes with 5-6 bars across the anal fin, may be confused with E. pusillus, E. tutuilae or E. ventermaculus. Enneapterygius pusillus and E. ventermaculus have distinctly U-shaped supratemporal sensory canals compared with a crescent in E. philippinus, and E. tutuilae has a high first dorsal fin, whereas in E. philippinus it is lower than the second. Enneapterygius philippinus also has 15 pectoral-fin rays vs. 14 for E. tutuilae. REMARKS. Snyder correctly recognised E. pardochir as the female of E. tusitalae (1912:518) but did not formally synonymise the two species. Fowler (1958) synonymised E. tusitalae and E. pardochir with E. philippinus, but did not state whether he recognised Jordan & Seale’s species as male and female of the same species. Schultz (1960) did, but neither of these authors state whether they examined Günther’s types. Günther’s description of E. minutus is brief in the extreme and merely notes that the two specimens on which the description is based were reddish-white with indistinct brown transverse bands on caudal and pectoral fins. His illustration of the species does show that the first dorsal fin is lower than the second (ibid: 213d). Fricke (1994) did examine Günther’s types and found that they agreed with specimens of E. pardochir, E. tusitalae, Tripterygion callionymi (Weber, 1909) and E. punctulatus (Herre, 1935) and synonymised the four species with E. minutus. Fricke (1997: 279-281) has a detailed analysis of the nomenclatural complications of the species and synonymised E. minutus with E. philippinus. Schultz’s inclusion of E. tutuilae Jordan & Seale as the same species as E. minutus is incorrect. The pigmentation of E. tutuilae is quite different: the species has a clearly banded anal fin, black mark in the margin of the second dorsal fin, and first dorsal fin distinctly higher that the second. MATERIAL EXAMINED. Pacific Ocean: New Hebrides: AMS I6317 (3); AMS I6418 (3); AMS I6451 (4); AMS I6582-3 (3). Lizard Island, Great Barrier Reef: ROM 72516 (21.7 & 21.9 mm). Abaiang Atoll, Gilbert & Ellis Islands: AMS I18049-011 (5). Indian Ocean: Comoro Ids: CAS 33458 (6: 17.4-21.5 mm). Mauritius: SAIAB 30163 (18.0 mm). Mahé, Seychelles: SAIAB 30155 (19.0 & 19.4 mm). Nosi Bé, Madagascar: AMS I28108 (4). Shimoni, Kenya: SAIAB 30158 (3: 15.2-17.8 mm). Ibo Island, Moçambique: SAIAB 30162 (6: 14.0-20.9 mm) & SAIAB 30157 (6: 16.8-21.3 mm). Pinda, Mocambique: SAIAB 30161 (17.7 & 20.0 mm). Cabo Delgado, Moçambique: SAIAB 30156 (12: 16.5-20.2 mm) & SAIAB 30159 (19.2 & 19.8 mm). Tekomasi Island, Moçambique: SAIAB 30154 (18.1 & 20.0 mm) & SAIAB 30160 (3: 18.0-20.9 mm). Enneapterygius pusillus Rüppell Enneapterygius pusillus Rüppell, 1835: 2, pl. 1, fig. 2 (Massawa, Red Sea); Clark, 1980: 108, Figs. 3a & 17 (Red Sea); Holleman, 1986: 757, Figure 236.5, non Plate 1a, b 16 (KwaZulu-Natal); Randall, 1995: 31 (Gulf of Oman). DESCRIPTION. Dorsal fins III+XII-XIV+10-11 (usually III+XIII+10-11); anal fins I,20-21 (see Table 1 for counts of Red Sea and Oman specimens); pectoral fins usually 13-14: 3-4+3-4+6-7 (usually 4+3+6). Lateral line with 912 pored scales and 25-28 notched scales, notched segment starting second scale row below end of pored segment (Fig. 16b). Total lateral scales 29-30; transverse scales 2/4; mandibular pores 2+2+2 (Fig. 16c). Head 3.6-3.9, body depth 5.3-6.0 in SL; eye 3.0-3.4 in head length. Small (less than 25 mm SL), slender fishes. Nape, belly and bases of dorsal and anal fins naked. First dorsal fin distinctly higher than second in males and about equal in height in females; second dorsal fin 70% of body depth. Pelvic-fin rays united by membrane for about half length of shorter ray. Maxilla reaching vertical through anterior of eye. Orbital cirrus small and lobate; supratemporal sensory canal deeply U-shaped. Live colour. Body colour of males pale cream with 4 irregular dark bars; scales on upper half with scattered orange and yellow-green spots and edged with a thin, deep orange line. Lower half of body with series of up to 10 orange blotches interspersed with irregular, dark, silver-grey spots. Abdomen covered anteriorly with dark brown rosettes and posteriorly with pale red blotches; dark brown or black pre-anal spot - often V-shaped usually present. Top of head laced with subcutaneous green and cream, with red surface spots. Upper margin of eye and orbital cirrus speckled with crimson. Snout and lips pale yellow with orange spots; cheeks with yellow, red and black spots. Pectoral fins yellow distally, orange proximally, with large circular orange-red mark with green centre on fin base. Pelvic fin bases and surrounding skin heavily mottled with deep red and black, rays black for proximal half, distally with red stipples; membrane black. First dorsal fin with light red spots and some melanophores; second dorsal fin heavily marked with dark brown to black melanophores interspersed with red spots and with 3 silvery-transparent vertical bands; third dorsal fin pale orange to transparent. Anal fin with red and black spots, the melanophores tending to form 6 or more indistinct bars. Upper half of caudal fin pale orange, lower half pale yellowish-green. Females less intensely pigmented and lack the melanin of large males. (Note: the colour photograph in Holleman, 1986 (plate 116) is not of E. pusillus but of E. tutuilae.) Colour in alcohol. All colour except for the melanin disappears, leaving bodies a pale straw colour, males with irregular bars across body, black on lower portion of head, thorax and extending to area around base of pelvic fins and proximal third to half of fins; pre-anal black spot present. Second and third dorsal and pectoral fins with irregular dusky bands; anal fin with basal spots and irregular black bars across fin. Females have the basal spots and bars on the anal fin, but lack black on the head, thorax and pelvics and dark body bars, having more irregular markings. DISTRIBUTION (Fig. 17). Known from inshore continental waters from the Red Sea south to KwaZulu-Natal, South Africa, from the Persian Gulf and coast of Oman (Randall, 1995), to the southeastern coast of India (Gulf of Mannar - Lal Mohan, 1971). COMPARISONS. Male E .pusillus are distinctive with their very high first dorsal fin and black pelvic-fin base and proximal portion of the fins. Females from the Red Sea may be confused with the sympatric E. pallidus, but E. pusillus usually has fewer anal-fin rays than the latter species: 17-21 (usually 19), vs. 21-22 (usually 22) for E. pallidus. Another sympatric species which shares the high first dorsal fin of male E. pusillus is E. tutuilae (= E. altipinnis - see below), but that species lacks a U-shaped supratemporal sensory canal and has fewer notched scales in the lateral line: 20-23 vs. 25-28 for E. pusillus. MATERIAL EXAMINED. SAIAB 7937 (21.7 mm); SAIAB 7941 (21.6 & 24.5 mm); SAIAB 7942 (6: 19.5-22.5 mm); SAIAB 7940 (5: 17.3-22.9 mm), all from Sodwana Bay, South Africa. Enneapterygius tutuilae Jordan & Seale Enneapterygius tutuilae Jordan & Seale, 1906: 418-419, figure 99 (Pago Pago, Samoa); Fricke 1994: 285ff, figures. 58 & 59; Randall, Allen & Steene, 1997: 364. Enneapterygius altipinnis: Clark, 1980: 99-101, figs. 3b, 6c, 11 (Aqaba & Eritrea, Red Sea). Enneapterygius pusillus: (non-Rüppell, 1835): Holleman, 1986: 757, pl. 116, fig. 236.5 (Australia). DESCRIPTION. Dorsal fins III+XI-XII+9-10 (usually III+XII+10); anal fin I,16-18 (usually 17 rays); pectoral-fin rays 14, all simple. Lateral line with 11-12 (usually 12) pored scales and 20-23 (usually 22) notched scales, notched segment starting second scale row below end of the pored segment (Fig. 18b). Total lateral scales 29; transverse scales 2/4. Mandibular pores 2+1/2+2 (Fig. 18c). Head 3.3-3.6, body depth 4.4-5.3 in SL; eye 2.6-3.2 in head length. Body moderately elongate. Nape scaled; belly naked. First dorsal fin higher than second in males and about the same height in females, second dorsal fin about 70% of body depth. Pelvic-fin rays united by membrane for about half length of shorter ray; longer ray reaches to vent. Longest pectoral-fin ray reaches end of second dorsal fin. Supratemporal sensory canal crescent-shaped. Maxilla reaches vertical through anterior margin of pupil. Orbital cirrus moderate and lobate. Live colour. There are two colour forms, an apparently more common green form and a brown form recorded from Phuket, Thailand and Mauritius. Green form (from transparencies made by R. Winterbottom in the Comoro Islands, Fiji and on Escape Reef, Australia, and by G. Allen [in: Randall et al., 1997] ) as follows: males with bright green bodies with white blotches along mid-side, a white saddle below rear half of second dorsal fin and a white bar across body at end of third dorsal fin; anal-fin base with 6-8 black spots; black preanal spot present; basal anal-fin spots and pre-anal mark absent in most St Brandon Shoals specimens. Scale-pocket margins on upper half of body lined with red dots. Snout, lower half of head, chest and pectoral-fin bases with many red spots; top of head green. First dorsal fin whitish-green; second with many red spots anteriorly, with black spots on the membranes between 4th and 9th spines and white band posteriorly, the white continuing onto the body; third dorsal fin transparent with some red basally and a white band on the margin posteriorly. Caudal-fin rays lined with red, membranes transparent; anal fin with alternating blackish red and white bars; pelvic fins spotted with red; pectoral fin rays lined in red but with 3 white bands, membranes transparent. The brown form with brown and orange spots on body and head and with narrow dark bar across caudal peduncle; first dorsal fin orange and cream; second as for the green form, but with orange and brown replacing the red. The white bands of the green form are creamcoloured in the brown form; lower half of pectoral fins red. Females of both forms less brightly coloured than males. Colour in alcohol. Males and females straw-coloured with 6-8 black bars across anal fin, dark basal spots on anal fin, prominent black pre-anal mark, and black blotch on margin of the middle of second dorsal fin. Mature males sometimes with stippling along inner pelvic fin ray, on head, and show indistinct, irregular, dusky bars across body. Immature males and females often show little pigment except the basal spots on anal fin and some black on second dorsal fin. DISTRIBUTION (Fig. 17). Widely distributed throughout the Indo-West Pacific Ocean, from East Africa to Japan and Samoa. COMPARISONS. Enneapterygius tutuilae, though similar to E. pusillus with its high first dorsal fin, has more prominent anal-fin bars, particularly the basal spots, which are absent in E. pusillus (see also remark above about St Brandon Shoals specimens), and lacks the deep U-shaped supratemporal sensory canal of E. pusillus. Enneapterygius ventermaculus shares the anal fin bars and basal spots with E. tutuilae, but also has a U-shaped supratemporal sensory canal and 21-25 notched lateral line scales, vs. 20-23 for E. tutuilae. Schultz (1960) considered E. tutuilae a synonym of E. minutus (= philippinus), together with E. tusitalae and E. pardochir. This is not so: E. tutuilae and E. philippinus are quite distinct in colour and form of the first dorsal fin. (See also under E. philippinus above). MATERIAL EXAMINED. Kenya: SAIAB 31521 (4:19.8-21.2 mm) Malindi. Mozambique: SAIAB 31522 (16.5 mm) Cabo Delagado; SAIAB 31515; (3: 15.0-18.6 mm) & SAIAB 15024 (17.4 mm), Baixo Pinda;. Indian Ocean: Comoro Ids: ROM 67490 (12.8 & 15.1 mm); ROM 67491 (13.6 & 15.1 mm); ROM 67493 (13.8 mm); ROM 67495 (6: 13.8-14.4 mm); ROM 67492 (13: 7.6-13.9 mm); ROM 67485 (14.6 mm); ROM 67486 (15.4 mm); ROM 67487 (11.6 &16.0 mm); ROM 67497 (8: 9.4-15.7 mm); ROM 67498 (15.6 mm); ROM 67494 (3: 11.8-14.1 mm); ROM 67499 (12.8&16.4 mm); ROM 67489 (12.9 & 14.9 mm); ROM 67500 (3: 14.8-15.0 mm); ROM 67496 (3: 10.1-15.0 mm); ROM 67484 (8: 14.3-15.8 mm); ROM 67488 (14.4 & 15.7 mm). Mauritius: SAIAB 31518 (14.7 mm); SAIAB 31519 (15.5 mm); SAIAB 31520 (14.8 & 15.2 mm). Chagos Archipelago: ROM 67467 (21.6 mm). St Brandon Shoals: SAIAB 31516 (16.0 mm); SAIAB 31517 (17.1 mm); USNM 357610 (18.5 mm); USNM 360012 (5: 17.5-22.1 mm); USNM 359861 (24: 14.6-19.4 mm); 359859 (16: 17.2-18.7 mm); USNM 359856 (4: 14.1-19.5 mm); USNM 359857 (16.9 & 18.5 mm).Pacific Ocean: Thailand: CAS 48185 (24: 11.1-17.4 mm); PMBC 17468 (3); PMBC17473 (5); PMBC 17475 (2). Japan: USNM 770713 (7: 9.3-20.4 mm). Enneapterygius ventermaculus Holleman Enneapterygius ventermaculus Holleman 1982: 123, fig. 7 (northern KwaZulu-Natal, South Africa); Randall 1995: 32, fig. 5 (Gulf of Oman). Enneapterygius nasima: Hoda 1983: 116ff, figs. 1, 2, 3 (Karachi coast, Pakistan). DESCRIPTION. Dorsal fin III+XI-XIII+8-10 (usually III+XII+10); anal fin I,17-20 (usually 19 rays); pectoral-fins rays 14: 1-3+4-6+6-8 (usually 2+5+7). Lateral line with 13-16 (usually 15) pored and 21-25 (usually 23) notched scales (see Table 1 for counts of Oman specimens), notched series starting second scale row below end of pored series (Fig. 19b); total lateral scales 31-34 (usually 32-33, but 35-36 for Oman specimens); transverse scales 2/6. Mandibular pores 3+1+3 (Fig. 19c). Head 3.4-4.0, body depth 4.8-5.2 in SL; eye 3.0-3.7 in head length. Body moderately robust; nape, base of first dorsal fin and belly naked. Mouth small and nearly horizontal, reaching vertical through anterior margin of orbit. Orbital cirrus moderate and lobate. Supratemporal sensory canal U-shaped, embracing first dorsal fin spine. First dorsal fin slightly higher than second in males, slightly lower in females; second dorsal fin about 70% of body depth. Pelvic-fin rays united by membrane for about half length of shorter ray, longer ray reaching as far as preanal mark. Live colour (also from Randall 1995). Body of males greenish, scale margins with brown stipples and orange edge, except ventrally; black pre-anal mark present, ranging from small spot to boomerang-shape half around vent; three white blotches along midside, first under end of pectoral fin, second below junction between second and third dorsal fins, third below end of third dorsal fin; caudal fin base with triangular dark area, apex anterior, base darkest, followed by narrow white band at base of caudal fin; head bottle-green above with orange-brown markings, darkest on sides below eyes, on thorax and to pelvic-fin bases and on pectoral-fin bases. First dorsal fin finely speckled with brown and orange, but without pigment on membrane between third spine and dorsum; second dorsal spines orange with alternating areas of orange and dense, dark brown stippling along fin margin and band of stipples along base; third dorsal fin banded with dark brown on the rays and orange stipples on membranes anteriorly; anal fin with 5-7 black broken bars with white and transparent areas between, and 57 black basal spots which continue subcutaneously; caudal fin rays greenish, upper with some brownish semi-bands, lowermost with black marks; pelvic fins yellow; pectoral fins with white and dark brown, lower rays with yellow, and two white spots between base of pectoral fin and base of pelvic fin, which may be largely obscured in darkly marked specimens. Females dorsally translucent pale green with orange margins to scales; no black or brown on ventral part of head and thorax, but small clusters of melanophores below midline more prominent than in males; a dark streak in front of each pelvic fin, another on side of thorax and a third from lower pectoral-fin base, latter two continuing a little below branchiostegal membrane. Colour in alcohol. The original description (Holleman 1982) noted no sexual dichromatism. However, Randall’s (1995) description and material available from Sur, Oman, shows males with densely clustered large melanophores in interorbital area, on head below eyes, opercles and lower portion of pectoral fin-bases, entire thorax, base of pelvic fins and anterior of belly, including proximal portion of fins themselves, the darkest a black band extending from lower pectoral fin bases across chest and pelvic fin bases. Body with irregular dusky markings that may form indistinct bars ventrally; caudal peduncle with triangular dark area, with densest stipples in band at base of caudal fin; dorsal fins with scattered melanophores, darkest between first two spines of first dorsal fin; anal fin with 5-7 irregular black bars, the colour on the rays only; pectoral fins irregularly marked with melanophores. Females irregularly pigmented with black and brown, tending to form 4-5 irregular H- or inverse Y-shaped bars along the sides. Prominent black preanal mark present, frequently in shape of triangle or boomerang-shape, apex anterior. Anal fin with 5-7 black basal spots which continue as irregular bars across the fin. Caudal and pectoral fins irregularly barred. DISTRIBUTION (Fig. 17). Aden, Oman, Pakistan and east coast of South Africa from northern KwaZulu-Natal to Tshani, Eastern Cape Province, an apparently anti-tropical distribution. COMPARISONS. Hoda (1983) differentiated Enneapterygius nasimae from E. ventermaculus on the basis of colour pattern; there is little to distinguish the two species morphometrically, the counts of E. nasimae falling within the range of those for E. ventermaculus. At the time of description the colour of E. ventermaculus was not known, but specimens from Pakistan were included in the type series (Holleman, 1982: 125). Fresh material subsequently available to me and Randall’s description and photograph of E. ventermaculus (1995: 32, fig. 5) from the coast of Oman agree with Hoda’s description of the colour of E. nasimae. Hoda’s only distinction is that E. nasimae has more regular blotches along the side than E. ventermaculus, particularly that on the caudal peduncle. His illustration (figs. 3a & b) suggests the triangular caudal mark as described by Randall. The species is distinctly not the same as E. hollemani (the differences are dealt with in detail under that species) and the E. nasimae is thus considered a junior synonym of E. ventermaculus. (See also under E. hollemani and E. tutuilae.) MATERIAL EXAMINED additional to that listed in Holleman 1982. Oman - ROM 57548 (22.0 mm); ROM 57549 (11.2 & 20.0 mm); ROM 57537 (3: 18.5-19.2 mm); ROM 57545 (4: 16.6-20.0 mm); ROM 57546 (3: 12.0-19.9 mm), all from Sur. South Africa - SAIAB 40336 (3: 21.0-27.6 mm) & SAIAB 40427 (25.8 & 26.6 mm), Park Rynie; SAIAB 40506 (26.7 mm), SAIAB 60242 (28.7 mm), SAIAB 60303 (6: 17.3-29.0 mm), Aliwal Shoal; SAIAB 53511 (31.0 mm), Tshani, Transkei; SAIAB 43242 (20.0 & 21.5 mm), Sodwana. COMMENTS ON INDIAN AND SRI LANKAN SPECIESThe most recent publications on tripterygiid fishes from the coasts of India and Sri Lanka are those of Lal Mohan (1968, 1971) and Hoda (1983). From those we know that at least two species of Enneapterygius are found: E. fasciatus and E. ventermaculus (E. nasimae). Fricke & Randall (1992) record E elegans and E. obscurus from the Maldives. It stands to reason that E. abeli, E. clarkae, E. gruschkai, E. minutus and, perhaps, E. tutuilae also occur in the Laccadive Islands and the islands around Sri Lanka. DISCUSSION ON COLOUR PATTERNSWith the exception of the New Zealand and southern Australian species, there is still little record of colour patterns of tripterygiids or information about associated behaviour patterns. While the number of colour photographs of species is increasing, including good underwater photographs by J. Randall, G. Allen, H. Debelius, R. Kuiter, H. Masuda, S. Dewa and others, few of the probably more than 150 species have been photographed or illustrated, and of only a handful of species are there photographs that show changes in colour patterns with growth, sexual dichromatism and the nuptial/spawning colours of males. While most species of Enneapterygius show sexual dichromatism, it is not certain that all do. This assumption was made in describing Enneapterygius ventermaculus (Holleman, 1982) and was found to be incorrect when specimens from Oman were examined. Thresher (1984) records that during spawning male tripterygiids generally develop some combination of black and/or red on the head, body and caudal fin. Zander & Heymer (1976) and Wirtz (1978:143) observed a black head in territorial males of all three Mediterranean species of Tripterygion, and both Shiogake & Dotsu (1973: 36) and Masuda & Kobayashi (1994: 310) figure a male Enneapterygius etheostomus in nuptial colours where nearly the entire body is black. While in all Enneapterygius spp. (probably in all tripterygiids) males are more brightly and/or darkly coloured than females, and a black or partially black head, thorax, abdomen and prepectoral area is not uncommon in males (E. abeli, E. elegans, E. minutus, E. pusillus, E. tutuilae and E. ventermaculus), it is not known whether the black coloration is sexual dichromatism or whether it only represents nuptual/territorial coloration, as it does in Tripterygion species. Randall (1993) suggests that the change in colour of large males of Helcogramma vulcana (and other species of tripterygiids) to entirely black does not seem to be associated with the attainment of sexual maturity, because pale individuals with dark bars may have large testes. One would not expect a fish to produce such an extreme amount of melanin for a brief spawning period, only to have it disappear after spawning. He records a more likely Jeffrey Williams’ hypothesis that melanistic males may be dominant in the local population (Randall 1993:31). This would seem to be in agreement with Wirtz’s observations (1978:143) Furthermore, in collections of Helcogramma obtusirostre from tide pools in KwaZulu-Natal, South Africa, where the species is abundant, few males are almost entirely black, while most are darker versions of females, an observation that may also corroborate Williams’ hypothesis. This degree of melanism has only been observed in males of Enneapterygius etheostomus (Masuda & Kobayashi 1994:310), E. kermadecensis, E. minutus and E. niger in which the males are very extensively covered with melanophores. Melanophore patterns may be of further value, as indicators of relationship. Fricke (1994:174) distinguishes six species groups for Australian and southwestern Pacific species of Enneapterygius, based on the length of the pored (anterior) portion of the lateral line, the amount of melanin in males and the height of the first dorsal fin, in that order, arbitrarily considering 12 or fewer pored scales as a short anterior lateral line and 13 or more as a long anterior lateral line. While this may be useful in a key, to use the character to define species groups implies phylogenetic relationship, for which Fricke offers no other substantiation. In Helcogramma there are examples of colour pattern indicating relationship as, for example, the H. fuscopinna species group as defined by Williams & McCormick (1990). The distinctive caudal peduncular bars of certain Enneapterygius species (E. clarkae, E. destai, E. elegans and E. gruschkai) may be indicative of relationship, as may be the body bars of E. clarkae, E. gruschkai and E. melanospilos, or the internally extending basal anal fin spots of E. hollemani, E. kosiensis, E. tutuilae and E. ventermaculus. Certainly the form of the supratemporal sensory canal with its associated osteological features, which may have phylogenetic implications, must be taken into consideration (Holleman, 1982). However, until study of the entire genus is undertaken these issues must remain speculative. ACKNOWLEDGEMENTS I am most grateful for the loan of material from the institutions listed, and particularly to Vic Springer and Rick Winterbottom. Jeff Williams of the Smithsonian Institution kindly provided data on Red Sea type specimens and Daniel Golani provided some of the type material of E. obscurus. Jeff Williams and Kendall Clements reviewed this paper and provided valuable comment. I also thank the Director of the South African Institute of Aquatic Biodiversity, Prof. Paul Skelton, for providing laboratory space and making the facilities of the Institute available for my use. LITERATURE CITED
© 2005 The South African Institute for Aquatic Biodiversity, Grahamstown, South Africa The following images related to this document are available:Photo images[sm05002f17.jpg] [sm05002f10.jpg] [sm05002f13.jpg] [sm05002f11.jpg] [sm05002f8.jpg] [sm05002f4.jpg] [sm05002f2.jpg] [sm05002f16.jpg] [sm05002f5.jpg] [sm05002f19.jpg] [sm05002f12.jpg] [sm05002t1.jpg] [sm05002f14.jpg] [sm05002p2.jpg] [sm05002f1.jpg] [sm05002p1.jpg] [sm05002f15.jpg] [sm05002f18.jpg] [sm05002p1a.jpg] [sm05002f7.jpg] [sm05002f3.jpg] [sm05002f6.jpg] [sm05002f9.jpg] [sm05002p2a.jpg] |
| |||||||||

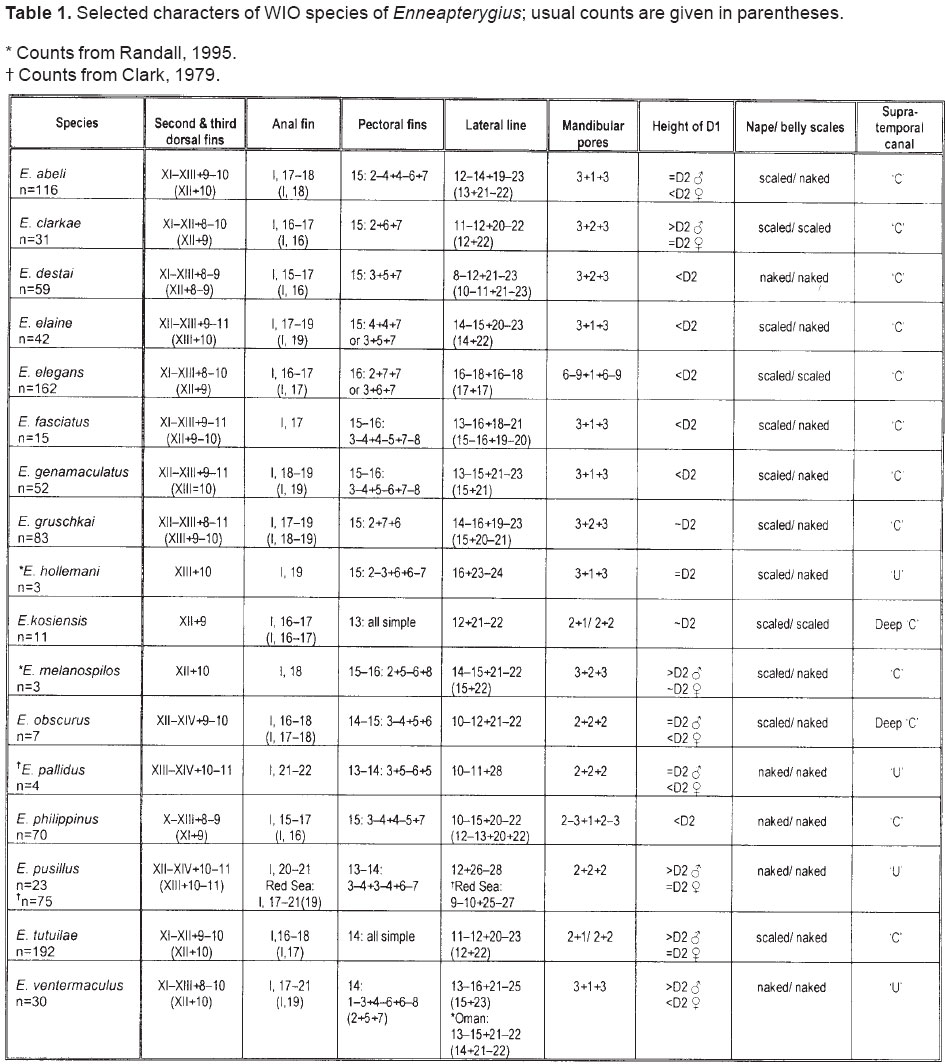{kind=link}
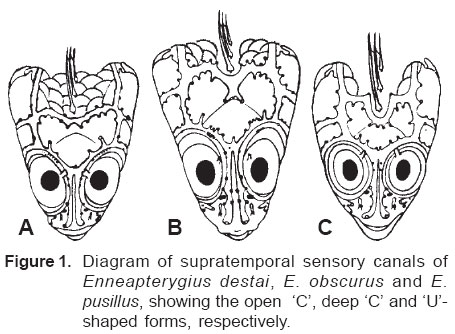{kind=link}
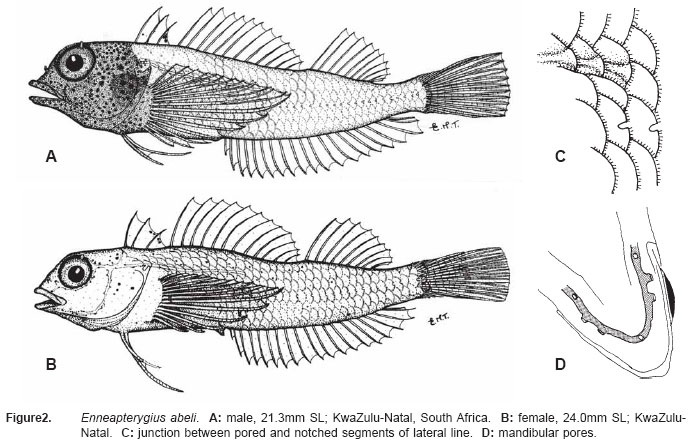{kind=link}
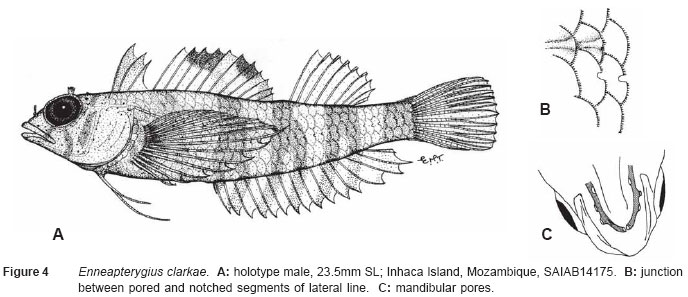{kind=link}
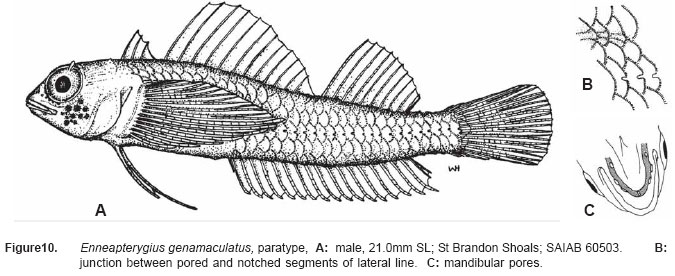{kind=link}
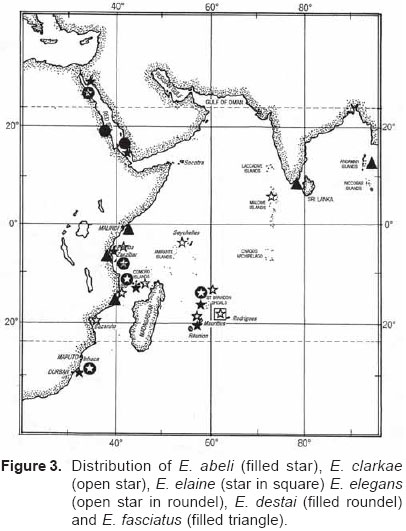{kind=link}
{kind=link}
{kind=link}
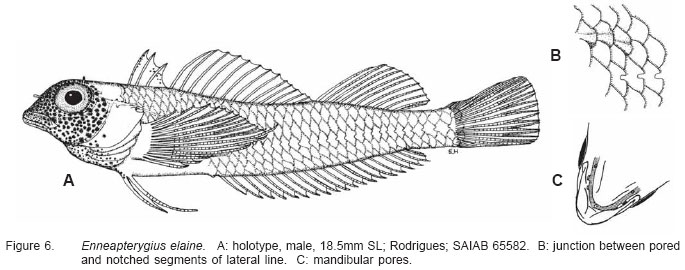{kind=link}
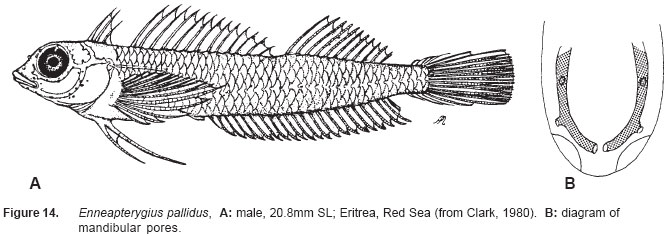{kind=link}
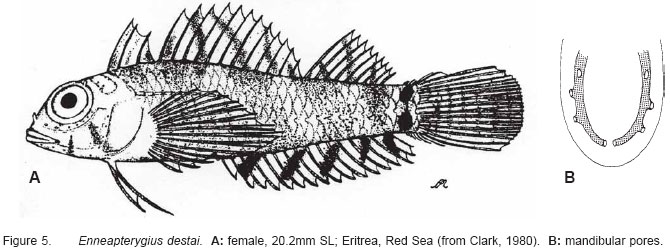{kind=link}
{kind=link}
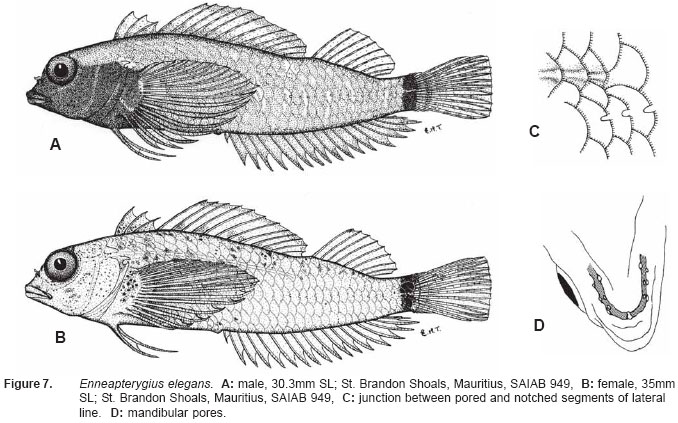{kind=link}
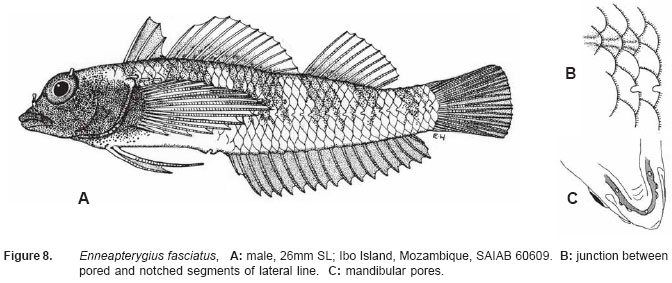{kind=link}
{kind=link}
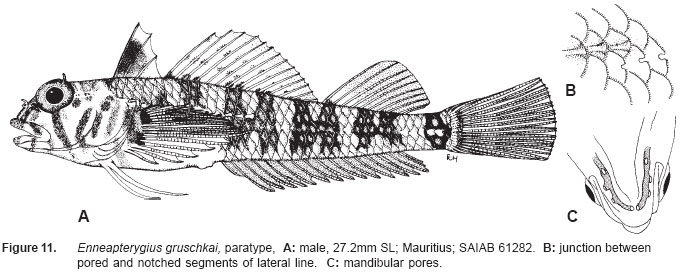{kind=link}
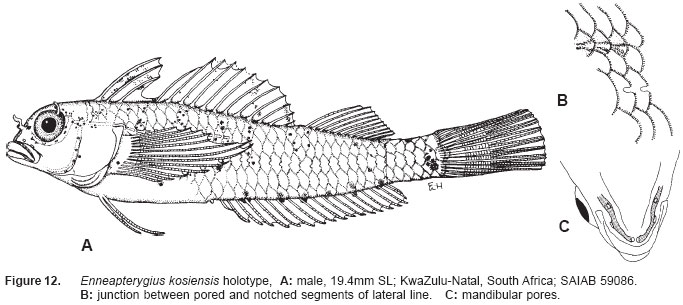{kind=link}
{kind=link}
{kind=link}
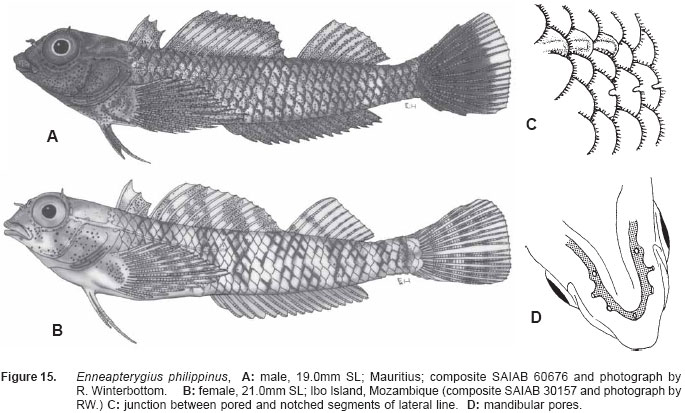{kind=link}
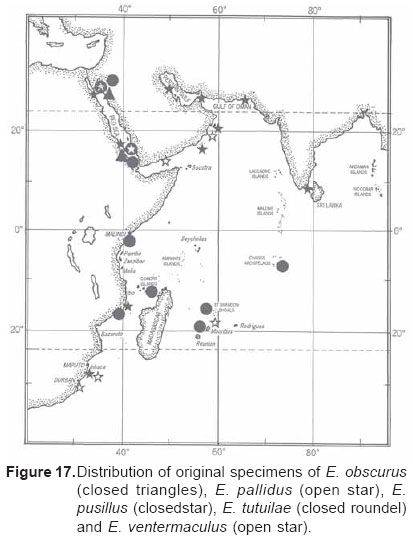{kind=link}
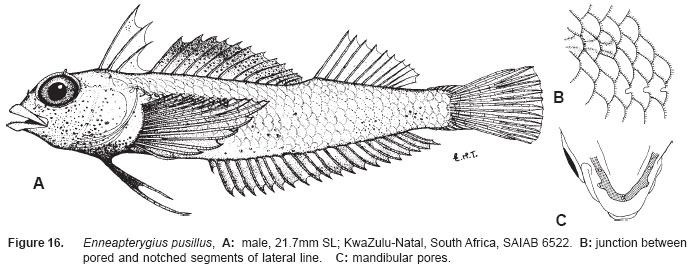{kind=link}
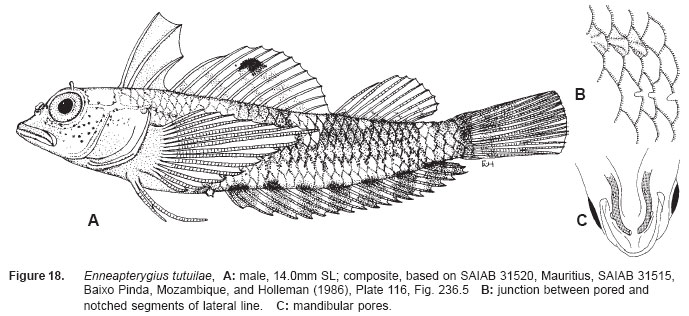{kind=link}
{kind=link}