 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
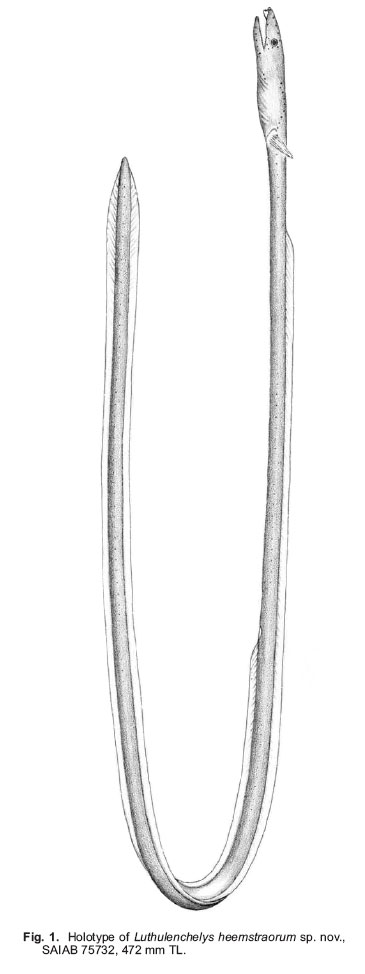
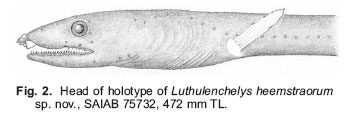
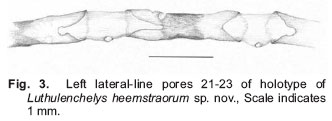
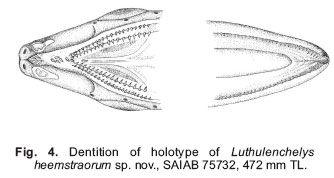
Smithiana, Publications in Aquatic Biodiversity, Bulletin 7, May, 2007, pp. 3-7 Luthulenchelys heemstraorum, a new genus and species of snake eel (Anguilliformes: Ophichthidae) from KwaZulu-Natal, with comments on Ophichthus rutidoderma (Bleeker, 1853) and its synonyms John E. McCosker California Academy of Sciences, San Francisco, California 94118, USA e-mail: Jmccosker@calacademy.org (Received 17 September 2006; accepted 2 January 2007) Code Number: sm07001 ABSTRACT Luthulenchelys heemstraorum genus and species novum, subfamily Ophichthinae, tribe Ophichthini, is described from a 472 mm eel trawled off Durban, KwaZulu-Natal, South Africa, in 450–460 m. Luthulenchelys differs from all known ophichthids in having the following suite of characters: an extremely elongate body, long tail, dorsal-fin origin in anterior trunk region, an elongate pectoral fin, posterior eye/jaw location, blunt snout, posterior nostril within upper lip, slender dentition, a single vomerine tooth, uniquely developed lateral-line ossicles, and five gill arches, with a very reduced fifth ceratobranchial. Several changes in taxonomy are proposed: Ophisurus lumbricoides Bleeker 1853, Ophisurus rutidoderma toides Bleeker 1853, and Ophichthus derbyensis Whitley 1941 are junior synonyms of Ophichthus rutidoderma (Bleeker 1853); Sphagebranchus lumbricoides Bleeker 1864 is a species of Yirrkala. KEY WORDS: Ophichthidae, Luthulenchelys heemstraorum gen. & sp. nov., KwaZulu-Natal, South Africa, Ophichthus rutidoderma, Ophichthus rutidodermatoides, Ophichthus derbyensis, Yirrkala lumbricoides INTRODUCTION The snake eel fauna of the Western Indian Ocean is rich, diverse and poorly known. J.L.B. Smith (1962) included 55 species in his monograph of the Western Indian Ocean and Red Sea ophicthids. McCosker and Castle’s (1986) account of the southern African species (from northern Namibia to Mozambique) treated 29 species and made several changes in taxonomy. In a forthcoming volume on coastal fishes of the Western Indian Ocean and Red Sea (Phillip Heemstra and Jack Randall, editors), I will report at least 70 species and 30 genera of ophichthids, including numerous changes in taxonomic status. In preparation for that work I have examined many of the recently collected specimens from South Africa and beyond that are deposited in the collections of the South African Institute for Aquatic Biology, the Bernice P. Bishop Museum, the United States National Museum of Natural History, the California Academy of Sciences, the British Museum of Natural History, and other institutions, and have discovered at least six species of ophichthids that appear to be undescribed, as well as a number of eels whose taxonomic history is as twisted as the state of their holotypes. Undoubtedly additional ophicthid species from this area remain to be discovered. The majority of the known undescribed species await description or explanation in a variety of generic revisions that are underway. One new species, however, is so distinct that it deserves a new genus as well, and in preparation for the publication of the Western Indian Ocean volume, I herein describe it and take pleasure in naming it after Phil and Elaine Heemstra in recognition of their prodigious efforts in this and previous regional works. MATERIALS AND METHODS The single specimen is deposited at the South African Institute for Aquatic Biodiversity (SAIAB), Grahamstown. Specimen measurements are straight-line, made either with a 300 mm ruler with 0.5 mm gradations (for total length, trunk length, and tail length) and recorded to the nearest 0.5 mm, or with dial calipers (all other measurements) and recorded to the nearest 0.1 mm. Body length comprises head and trunk lengths. Head length is measured from the snout tip to the postero-dorsal margin of the gill opening; trunk length is taken from the gill opening to mid-anus; maximum body depth does not include the median fins. Head-pore terminology follows McCosker et al. (1989: 257), in which supraorbital pores include the ethmoidal pore + pores in supraorbital canal, i.e. 1 + 3, and the infraorbital pores include pores along the upper jaw + those in the vertical canal behind the eye (“postorbital pores”), i.e. 4 + 2, as the last pore included along the upper jaw is frequently part of the postorbital series. Gill arch and lateral-line ossicle examination was accomplished after removal and clearing and counterstaining with alcian blue and alizarin red dyes (Dingerkus & Uhler 1977). Vertebral counts (which include the hypural) were taken from radiographs. Radiographic techniques are described in (Böhlke 1989). The mean vertebral formula (MVF) is expressed as the average of predorsal, preanal, and total vertebrae (Böhlke, 1982). Institutional abbreviations follow (Leviton et al., 1985). Luthulenchelys gen. nov. Type species: Luthulenchelys heemstraorum sp. nov. DIAGNOSIS. An elongate ophichthid, subfamily Ophichthinae, tribe Ophichthini (sensu McCosker 1977), with tail much longer than head andtrunk; median fins low; dorsal fin arising well behind pectoral fin; pectoral fin base arising above and occupying more than half of gill opening; gill openings lateral, elongate, nearly vertical and crescentic; eye moderately developed, its centre above posterior quarter of upper jaw, its posterior margin slightly in advance of rictus; jaws moderately developed, but not elongate; snout conical, tapering evenly, its tip rounded; underside of snout with a median sulcus exposing the teeth in advance of anterior nostril bases; anterior nostrils within short tubes; posterior nostrils a hole within upper lip, covered by a flap; teeth conical, erect, numerous and small, uniserial on mandible and biserial on maxillary, a single vomerine tooth; 2 preopercular pores; gill arches similar to those of Ophichthus (cf. Nelson 1966;McCosker 1977), however, fifth ceratobranchial is a thin, nearly ossified rod for anterior 80%, the remainder cartilaginous; upper pharyngeal tooth plates fused. Other characters those of the single species. ETYMOLOGY. Named Luthulenchelys in honour of Chief Albert John Mvumbi Luthuli of KwaZulu-Natal, Africa’s first winner of the Nobel Peace Prize and former President of the African National Congress; and enchelys, an ancient Greek word for eel, feminine. REMARKS. In general appearance, the type species of Luthulenchelys appears similar to some of the very elongate species of Ophichthus, such as O. rutidoderma and O. microcephalus. Luthulenchelys heemstraorum differs from them in the condition of its upper lip and associated posterior nostril, its large posterior eye (ending nearly above rictus, rather than in advance of it), its reduced vomerine dentition, its fifth ceratobranchial reduced to a minor cartilaginous rod (rather than an ossified structure), and in the condition of its lateral-line ossicles. Adaptations displayed by the new species such as the nearly uniform dark coloration, small and numerous teeth, fairly large eye, tumid snout (associated with burrowing in soft mud, rather than the sharp-pointed snouts and tails of shallow-water ophichthids that are adaptations for burrowing into sand), moderately developed cephalic pores, and poorly developed median fins are shared by other deep-dwelling ophichthids (cf. McCosker et al. 1989; McCosker 1999). Those similarities and adaptations are so apparent that upon first glance the new species appears very similar to several deep-dwelling Atlantic myrophine ophichthids (McCosker, 1989), such as Asarcenchelys longimanus McCosker 1985, Mixomyrophis pusillipinna McCosker 1985 and Pseudomyrophis nimius Böhlke 1960, whose ancestry is very different to that of the new species. Luthulenchelys heemstraorum sp. nov. Holotype: SAIAB 75732, 472 mm TL, sex undetermined (gonads not apparent), from KwaZulu-Natal, South Africa, off Durban (29°57’14”S, 31°16’23”E), 450–460 m, collected by Grant van der Westhuizen aboard F/V Ocean Spray using an otter trawl, field no. ORI 180-3/5, on 16 November 2004. DIAGNOSIS. An elongate species of ophichthine with: tail 62%, head 7.2%, and body depth at gill opening 1.3% of total length; dorsal-fin origin nearly 2 head lengths behind pectoral-fin tips; pectoral fin elongate; posterior nostril a hole within upper lip, covered by a flap that extends below edge of lip; no barbels on upper lip and a no slit at posterior nostril; pores small but conspicuous, SO 1 + 4, IO 4 + 2, POM 5 + 2; teeth small and conical, biserial on upper jaw, uniserial on lower jaw, a single vomerine tooth; coloration uniform grey-black; vertebral formula 20/61/165. Counts and measurements of the holotype (in mm). Total length 472; head 34.0; trunk 144; tail 294; predorsal distance 64; pectoral-fin length 8.25; pectoral base 2.4; body depth ~6 at gill openings; body width ~6 at gill openings; body depth ~5 at anus; body width ~5 at anus; snout 7.5; tip of snout to rictus 11.2; snout overhang 3.5; eye diameter 3.0; interorbital width 3.5; gill opening height 3.1; isthmus width 2.6. DESCRIPTION. Body very elongate, trunk subcircular, tail laterally compressed; body depth at gill openings 79 in TL. Branchial basket moderately expanded; 17 pairs of overlapping branchiostegal rays visible by radiograph. Head 4.2 in trunk. Head and trunk 2.7 and head 14 in TL. Snout rounded, moderately acute when viewed from above; a short groove bisecting underside of snout nearly to tip of upper jaw. A pair of small bumps on snout flanking dorsal midline in advance of anterior nostril base (their appearance possibly exaggerated by condition of specimen). Lower jaw included, its tip reaching well beyond base of anteriornostril tubes. Upper jaw not elongated, rictus immediately behind a vertical line at posterior margin of eye. Eye fairly large, 3.7 in upper jaw and 11.3 in head. Anterior nostrils tubular, extending ventrolaterally from snout at ~45° from horizontal, reaching below upper lip and beyond tip of chin. Posterior nostrils an elongate opening within upper lip, not visible externally, covered by a flap (expanded edge of lip) that extends below edge of mouth and lacks a vertical slit. No barbels along upper lip. Dorsal-fin origin well behind pectoral fin about a head length into trunk length. Median fins low but obvious, ending a little more than eye diameter before bluntly pointed and laterally compressed tail tip. Pectoral fins elongate. Head pores small but apparent (Fig. 2). Single median interorbital and temporal pores. Supraorbital pores 1 + 4, infraorbital pores 4 + 2, lower jaw pores 5, preopercular pores 2, supratemporal pores 3. Lateralline pores apparent only in anterior trunk region; 9 before gill opening in a high-arching sequence, the remainder difficult to discern due to condition of specimen. Lateral-line canal unique in appearance (Fig. 3) among the Ophichthini, exhibiting horizontal tubes indented at each end; pores in tail region appear as prominent sharp spikes as seen from above (these are lateral processes of the vertebrae as evidenced by radiography, perhaps exaggerated by the desiccated condition of the specimen), reaching nearly to tail tip. Teeth (Fig. 4) small, conical, slightly recurved; biserial in upper jaw and uniserial on mandible. An intermaxillary rosette of 4, followed by a gap, then a diamond of 4, followed by a single vomerine teeth. Maxillary tooth rows linear, separated by a narrow gap; inner row of ~16–17 larger teeth, flanked medially by ~22–24 smaller teeth in outer row. Lower-jaw teeth uniserial, ~28–30 descending in size to become very small posteriorly. Gill arches removed, cleared and counterstained. Gill arches developed, similar to those of Ophichthus (cf. McCosker 1977: 32) except for fifth ceratobranchial condition; first basibranchial ossified, all others cartilaginous; hypobranchials 1–2 ossified, 3–4 cartilaginous; epibranchials 1–4 ossified; infrapharyngobranchials 2–3 ossified; fifth ceratobranchial very reduced, a thin nearly ossified rod for anterior 80%, the remainder cartilaginous; upper pharyngeal tooth plates fused, containing 2–4 rows of conical recurved teeth; lower pharyngeal tooth plates contain 2–3 rows of 6–14 conical recurved teeth. Colour in ethanol: uniform grey-black; throat, snout and chin slightly darker; median and pectoral fins pale; anterior nostrils, inside of mouth, anal region and peritoneum pale. A photograph of the dead specimen taken soon after its capture indicates that in life it was uniform dark brown. ETYMOLOGY. Named in honour of Phil and Elaine Heemstra in recognition of their efforts to understand, illustrate, and explain the fishes of the Indian Ocean to scientists and the general public. DISTRIBUTION. Known only from the type specimen, collected off Durban in 450–460 m depth. REMARKS AND COMPARISONS. The characteristics of the new species are those of the monotypic genus. The new species is easily separable from any known ophichthid on the basis of its eye size and location, its extremely reduced vomerine dentition, its body elongation, and other characters unique to the genus. Comparisons with species of Ophichthus that are similar in appearance are described in the following section. COMMENTS ON OPHICHTHUS RUTIDODERMA During my initial attempts to identify the specimen herein described as Luthulenchelys heemstraorum, I examined the extremely elongate species of Ophichthus, O. microcephalus and O. rutidoderma. Ophichthys microcephalus Day 1878 was described from Malabar, India, and is currently known only from Day’s description and an extant 625 mm TL syntype in the Australian Museum (B.7843). (Ophichthus microcephalus may ultimately deserve relocation to another genus, however, the extant syntype is somewhat desiccated and its head is damaged, which would disallow an adequate generic decision.) I compared that syntype to the holotype of Luthulenchelys heemstraorum and found them to differ in numerous characters: the teeth of O. microcephalus are more numerous, more closely-set, and stouter than those of L. heemstraorum; the snout is shorter and sharper, the body less elongate, the eye is comparatively smaller and located more anteriorly, and O. microcephalus has more vertebrae than L. heemstraorum (12/69/209 vs. 20/61/165); and its lateral-line ossicles are more typical of the Ophichthus condition (McCosker 1977) than are those of L. heemstraorum. I also compared the shallow-water Indonesian species Ophichthus rutidoderma (Bleeker 1853) (treated by Kaup 1856: 18, as Pisoodonophis rutidoderma, later emended to Ophichthys rhytidoderma by Günther 1870: 63) to Luthulenchelys heemstraorum. It has a similar dorsal-fin origin (above the 16th vertebra) and biserial dentition, but the snout is shorter and sharper, its eye is smaller and located more anteriorly than that of L. heemstraorum, its lateral-line ossicles are more typical of the Ophichthus condition (McCosker 1977), and it has more total vertebrae (195-199 vs. 165). While examining the type (BMNH 1867.11.28:226, 94.5 cm TL) of Ophisurus rutidoderma Bleeker (1853: 30) at the British Museum, I attempted to decipher its identity and that of related Bleeker types. I ultimately concluded that Ophisurus rutidoderma toides Bleeker (1853: 31) [treated by Kaup 1856: 18, as Pisoodonophis rutidermatoides (sic.), later emended to Ophichthys rhytidodermatoides by Günther 1870: 62] and Ophisurus lumbricoides Bleeker (1853: 32) are junior synonyms of O. rutidoderma . The types of O. rutidodermatoides (BMNH 1867.11.28:292, 61 cm TL) and O. lumbricoides (BMNH 1867.11.28:300, 41.5 cm TL) are in poor condition, however, that of O. rutidoderma is in fair condition. Eschmeyer (1998: 1491) cited O. rutidoderma and O. rutidodermatoides as valid species whose unique holotypes were “whereabouts unknown.” My examination of those Bleeker specimens in the British Museum demonstrated that the length and locality data were appropriate, and I therefore identify those specimens, with confidence, as the holotypes. Eschmeyer (1998: 951) listed O. lumbricoides as a valid species of Yirrkala, and also suggested that the whereabouts of the unique holotype was unknown. It is in fact the British Museum specimen, as evidenced by the length of the specimen and additional records in the collection; Eschmeyer’s assumption of its validity was based upon McCosker and Castle’s (1986: 185)mistaken referral of Ophisurus lumbricoides Bleeker 1853, rather than Sphagebranchus lumbricoides Bleeker 1864, to Yirrkala in their treatment. I have also examined the holotype (AM.I.840, 258 mm TL) of Ophichthus derbyensis Whitley 1941 described from a specimen from Derby,Western Australia, and found it to be a synonym of Ophichthus rutidoderma. It has the same proportions, meristics, and dental condition as that of Bleeker’s type specimens. The vertebral formulae of the four holotypes are as follows: O. derbyensis 14/61/195; O. lumbricoides 16/64/199; O. rutidoderma 16/61/197; and O. rutidodermatoides ?/63/197. In summary, the twisted tale of these elongate ophichthids is as follows: Ophisurus lumbricoides Bleeker 1853, Ophisurus rutidodermatoides Bleeker 1853, and Ophichthus derbyensis Whitley 1941 are herein considered to be junior synonyms of Ophichthus rutidoderma (Bleeker 1853), and Sphagebranchus lumbricoides Bleeker 1864 is a valid species of Yirrkala. ACKNOWLEDGEMENTS My list of acknowledgments could easily exceed the length of the elongate holotype of the new species. I sincerely thank the following for their assistance: Grant van der Westhuizen and Sean Fennessy of theSouth African Oceanographic Research Institute (ORI) for collecting and photographing the type specimen; Beth Herd Guy for preparing the illustrations; Mysi Hoang (CAS) for clearing and staining the gill arches and skin segment; William N. Eschmeyer (CAS) for advice concerning nomenclature; the research and curatorial staff members of the Australian Museum (AMS), the British Museum of Natural History (BMNH), the Bernice P. Bishop Museum (BPBM), the California Academy of Sciences (CAS), the South African Institute for Aquatic Biodiversity (SAIAB), the United States National Museum of Natural History (USNM), and the Universiteit van Amsterdam Zoologisch Museum (ZMA) for their assistance with specimens; Phil Heemstra and the late Margaret Mary Smith for their advice and assistance during my visits to SAIAB; Tomio Iwamoto (CAS) for reading a draft of this manuscript; and Don Linker and the Jewish Community Endowment Fund for their financial support of the artist. LITERATURE CITED
© 2007 The South African Institute for Aquatic Biodiversity, Grahamstown, South Africa The following images related to this document are available:Photo images[sm07001f4.jpg] [sm07001f2.jpg] [sm07001f1.jpg] [sm07001f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}